Нейроны
Отдельные нервные клетки, или нейроны, выполняют свои функции не как изолированные единицы, подобно клеткам печени или почек. Работа 50 миллиардов (или около того) нейронов нашего мозга состоит в том, что они получают сигналы от каких-то других нервных клеток и передают их третьим.
Передающие и принимающие клетки объединены в нервные цепи или сети (см. рис. 26). Отдельный нейрон с дивергентной структурой (от лат. diverge — отклоняюсь) может посылать сигналы тысяче и даже большему числу других нейронов. Но чаще один такой нейрон соединяется всего лишь с несколькими определенными нейронами. Точно так же какой-либо нейрон может получать входную информацию от других нейронов с помощью одной, нескольких или многих входных связей, если на нем сходятся конвергентные пути (от лат. converge — приближаюсь, схожусь). Конечно, все зависит от того, какую именно клетку мы рассматриваем и в какую сеть она оказалась включенной в процессе развития. Вероятно, в каждый момент времени активна лишь небольшая часть путей, оканчивающихся на данном нейроне.
Действительные места соединения — специфические точки на поверхности нервных клеток, где происходит их контакт, — называются синапсами (synapsis; греч. «соприкосновение», «соединение») (см. рис. 26 и 27), а сам процесс передачи информации в этих местах — синаптической передачей. При взаимодействии нейронов с помощью синаптической передачи посылающая сигнал (пресинаптическая) клетка выделяет определенное вещество на рецепторную поверхность воспринимающего (постсинаптического) нейрона. Это вещество, называемое нейромедиатором, служит молекулярным посредником для передачи информации от передающей клетки к воспринимающей. Нейромедиатор замыкает цепь, осуществляя химическую передачу информации через синаптическую щель — структурный разрыв между передающей и воспринимающей клетками в месте синапса.
Особенности нервных клеток
Нейроны обладают рядом признаков, общих для всех клеток тела. Независимо от своего местонахождения и функций любой нейрон, как и всякая другая клетка, имеет плазматическую мембрану, определяющую границы индивидуальной клетки. Когда нейрон взаимодействует с другими нейронами или улавливает изменения в локальной среде, он делает это с помощью плазматической мембраны и заключенных в ней молекулярных механизмов.
Все, что находится внутри плазматической мембраны (кроме ядра), называется цитоплазмой. Здесь содержатся цитоплазматические органеллы, необходимые для существования нейрона и выполнения им своей работы (см. рис. 27 и 28). Митохондрии обеспечивают клетку энергией, используя сахар и кислород для синтеза специальных высокоэнергетических молекул, расходуемых клеткой по мере надобности. Микротрубочки — тонкие опорные структуры — помогают нейрону сохранять определенную форму. Сеть внутренних мембранных канальцев, с помощью которых клетка распределяет продукты, необходимые для ее функционирования, называется эндоплазматическим ретикуломом.
Существует два вида эндоплазматического ретикулума. Мембраны «шероховатого», или гранулярного, ретикулума усеяны рибосомами, необходимыми клетке для синтеза секретируемых ею белковых веществ. Обилие элементов шероховатого ретикулума в цитоплазме нейронов характеризует их как клетки с весьма интенсивной секреторной деятельностью. Белки, предназначенные только для внутриклеточного использования, синтезируются на многочисленных рибосомах, не прикрепленных к мембранам ретикулума, а находящихся в цитоплазме в свободном состоянии. Другой вид эндоплазматического ретикулума называют «гладким». Органеллы, построенные из мембран гладкого ретикулума, упаковывают продукты, предназначенные для секреции, в «мешочки» из таких мембран для последующего переноса их к поверхности клетки, где они выводятся наружу. Гладкий эндоплазматический ретикулум называют также аппаратом Гольджи, по имени итальянца Эмилио Гольджи, который впервые разработал метод окраски этой внутренней структуры, сделавший возможным ее микроскопическое изучение.

Камилло Гольджи (1844-1926). Фотография сделана в начале 1880-х годов, когда Гольджи был профессором университета в Павии. В 1906 году он разделил с Кахалом Нобелевскую премию по физиологии и медицине.

Сантьяго Рамон-и-Кахал (1852-1934). Поэт, художник и гистолог, обладавший поразительным творческим потенциалом, он преподавал в основном в Мадридском университете. Этот автопортрет он создал в 1920-х годах.
В центре цитоплазмы находится клеточное ядро. Здесь у нейронов, как и у всех клеток с ядрами, содержится генетическая информация, закодированная в химической структуре генов. В соответствии с этой информацией полностью сформированная клетка синтезирует специфические вещества, которые определяют форму, химизм и функции этой клетки. В отличие от большинства других клеток тела зрелые нейроны не могут делиться, и генетически обусловленные продукты любого нейрона должны обеспечивать сохранение и изменение его функций на протяжении всей его жизни.
Нейроны сильно различаются по своей форме, по связям, которые они образуют, и способам функционирования. Наиболее очевидное отличие нейронов от других клеток заключается в разнообразии их размеров и формы. Большинство клеток тела имеет шарообразную, кубическую или пластинчатую форму. Для нейронов же характерны неправильные очертания: у них имеются отростки, часто многочисленные и разветвленные. Эти отростки — живые «провода», с помощью которых образуются нейронные цепи. Нервная клетка имеет один главный отросток, называемый аксоном (греч. ax?n — ось), по которому она передает информацию следующей клетке в нейронной цепи. Если нейрон образует выходные связи с большим числом других клеток, его аксон многократно ветвится, чтобы сигналы могли дойти до каждой из них.
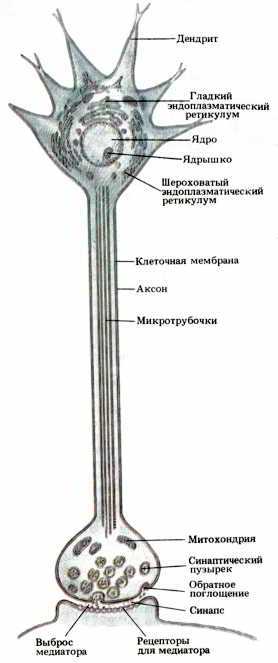
Рис. 28. Внутреннее строение типичного нейрона. Микротрубочки обеспечивают структурную жесткость, а также транспортировку материалов, синтезируемых в теле клетки и предназначенных для использования в окончании аксона (внизу). В этом окончании находятся синаптические пузырьки, содержащие медиатор, а также пузырьки, выполняющие иные функции. На поверхности постсинаптического дендрита показаны предполагаемые места рецепторов для медиатора (см. также рис. 29).
Другие отростки нейрона называются дендритами. Этот термин, происходящий от греческого слова dendron — «дерево», означает, что они имеют древовидную форму. На дендритах и на поверхности центральной части нейрона, окружающей ядро (и называемой перикарионом, или телом клетки), находятся входные синапсы, образуемые аксонами других нейронов. Благодаря этому каждый нейрон оказывается звеном той или иной нейронной сети.
В разных участках цитоплазмы нейрона содержатся различные наборы специальных молекулярных продуктов и органелл. Шероховатый эндоплазматический ретикулум и свободные рибосомы обнаружены только в цитоплазме тела клетки и в дендритах. В аксонах эти органеллы отсутствуют, и поэтому синтез белка здесь невозможен. Окончания аксонов содержат органеллы, называемые синаптическими пузырьками, в которых находятся молекулы медиатора, выделяемого нейроном. Полагают, что каждый синаптический пузырек несет в себе тысячи молекул вещества, которое используется нейроном для передачи сигналов другим нейронам (см. рис. 29).
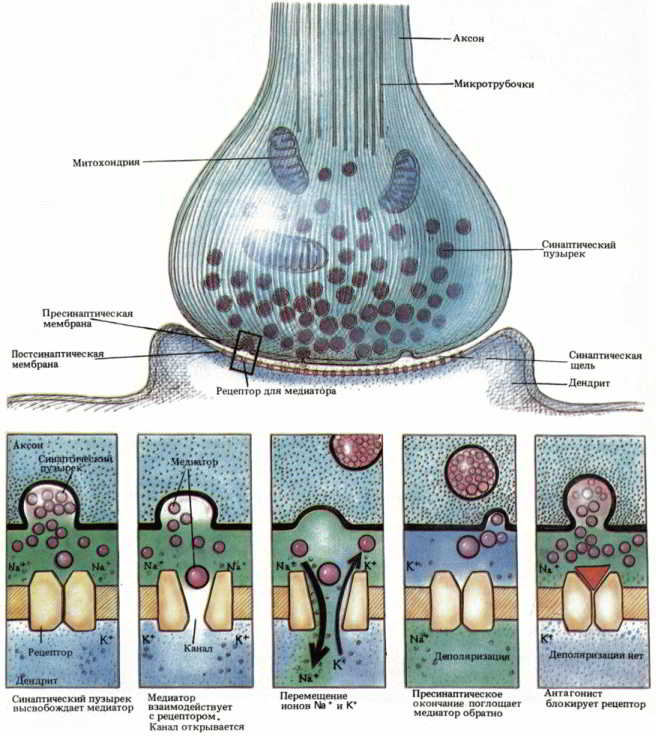
Рис. 29. Схема выброса медиатора и процессов, происходящих в гипотетическом центральном синапсе.
Дендриты и аксоны сохраняют свою форму благодаря микротрубочкам, которые, по-видимому, играют также роль в передвижении синтезированных продуктов из центральной цитоплазмы к очень далеким от нее концам ветвящихся аксонов и дендритов. При методе окраски, разработанном Гольджи, используется металлическое серебро, которое связывается с микротрубочками и позволяет выявить форму изучаемой нервной клетки. В начале XX века испанский микроанатом Сантьяго Рамон-и-Кахал почти интуитивно применил этот метод для установления клеточной природы организации мозга и для классификации нейронов в соответствии с их уникальными и общими структурными особенностями.
Различные названия нейронов
Нейроны в зависимости от контекста могут называться по-разному. Иногда это может сбить с толку, но на самом деле это очень похоже на то, как мы называем себя или наших знакомых. Смотря по обстоятельствам, мы говорим об одной и той же девушке как о студентке, дочери, сестре, рыжеволосой красавице, пловчихе, любимой или члене семейства Смитов. Нейроны тоже получают столько ярлыков, сколько различных ролей они выполняют. Разные ученые использовали, вероятно, все достойные внимания свойства нейронов в качестве основы для их классификации.
Каждая уникальная структурная особенность того или иного нейрона отражает степень его специализации для выполнения определенных задач. Можно называть нейроны в соответствии с этими задачами, или функциями. Это один способ. Например, нервные клетки, объединенные в цепи, которые помогают нам воспринимать внешний мир или контролировать события, происходящие внутри нашего тела, именуются сенсорными (чувствительными) нейронами. Нейроны, объединенные в сети, вызывающие мышечные сокращения и, следовательно, движение тела, называются моторными или двигательными.
Положение нейрона в сети — другой важный критерий наименования. Нейроны, ближе всего расположенные к месту действия (будь то ощущаемый стимул или активируемая мышца), — это первичные сенсорные или моторные нейроны, или нейроны первого порядка. Далее следуют вторичные нейроны (нейроны второго порядка), затем третичные (третьего порядка) и т.д.
Регуляция нейронной активности
Способность нервной системы и мышц генерировать электрические потенциалы известна давно — со времен работ Гальвани в конце XVIII столетия. Однако наши знания о том, как возникает это биологическое электричество при функционировании нервной системы, основаны на исследованиях всего лишь 25-летней давности.
Все живые клетки обладают свойством «электрической полярности». Это означает, что по отношению к какой-то отдаленной и явно нейтральной точке (электрики называют ее «землей») внутренняя часть клетки испытывает относительный недостаток положительно заряженных частиц и поэтому, как мы говорим, отрицательно заряжена относительно наружной стороны клетки. Что же это за частицы, находящиеся внутри и вне клеток нашего тела?
Жидкости нашего тела — плазма, в которой плавают клетки крови, внеклеточная жидкость, заполняющая пространство между клетками различных органов, спинномозговая жидкость, находящаяся в желудочках мозга, — все это особые разновидности соленой воды. (Некоторые ученые, мыслящие историческими категориями, усматривают в этом следы того периода эволюции, когда все живые создания существовали в первичном океане.) Встречающиеся в природе соли обычно составлены из нескольких химических элементов — натрия, калия, кальция и магния, несущих положительные заряды в жидкостях тела, и хлорида, фосфата и остатков некоторых более сложных кислот, образуемых клетками и несущих отрицательный заряд. Заряженные молекулы или атомы именуются ионами.
Во внеклеточных пространствах положительные и отрицательные ионы распределены свободно и в равных количествах, так что они нейтрализуют друг друга. Внутри клеток, однако, относительный дефицит положительно заряженных ионов обусловливает общий отрицательный заряд. Этот отрицательный заряд возникает потому, что плазматическая мембрана проницаема не для всех солей в равной мере. Некоторые ионы, например К+, обычно проникают сквозь мембрану легче, чем другие, такие, как ионы натрия (Na+) или кальция (Ca2+). Внеклеточные жидкости содержат довольно много натрия и мало калия. Внутри клеток жидкости относительно бедны натрием и богаты калием, но общее содержание положительных ионов внутри клетки не вполне уравновешивает отрицательные заряды хлорида, фосфата и органических кислот цитоплазмы. Калий проходит через клеточную мембрану лучше других ионов и, по-видимому, весьма склонен выходить наружу, так как концентрация его внутри клеток намного выше, чем в окружающей их среде. Таким образом, распределение ионов и избирательность их перехода через полупроницаемую мембрану приводят к созданию отрицательного заряда внутри клеток.
В то время как описанные факторы ведут к установлению трансмембранной ионной полярности, другие биологические процессы способствуют ее поддержанию. Один из таких факторов — очень эффективные ионные насосы, которые существуют в плазматической мембране и получают энергию от митохондрий. Такие насосы «откачивают» ионы натрия, поступающие в клетку с молекулами воды или сахара.
«Электрически возбудимые» клетки, подобные нейронам, обладают способностью регулировать свой внутренний отрицательный потенциал. При воздействии некоторых веществ в «возбуждающих» синапсах свойства плазматической мембраны постсинаптического нейрона изменяются. Внутренность клетки начинает терять свой отрицательный заряд, и натрий не встречает больше препятствий для перехода внутрь через мембрану. Действительно, после проникновения в клетку некоторого количества натрия переход натрия и других положительных ионов (кальция и калия) внутрь клетки, т.е. деполяризация, во время краткого периода возбуждения протекает столь успешно, что внутренность нейрона менее чем на 1/1000 секунды становится заряженной положительно. Этот переход от обычного отрицательного состояния содержимого клетки к кратковременному положительному называют потенциалом действия или нервным импульсом. Положительное состояние длится так недолго потому, что реакция возбуждения (повышенное поступление в клетку натрия) носит саморегулируемый характер. Присутствие повышенных количеств натрия и кальция в свою очередь ускоряет эвакуацию калия, по мере того как ослабевает действие возбуждающего импульса. Нейрон быстро восстанавливает электрохимическое равновесие и возвращается к состоянию с отрицательным потенциалом внутри до следующего сигнала.
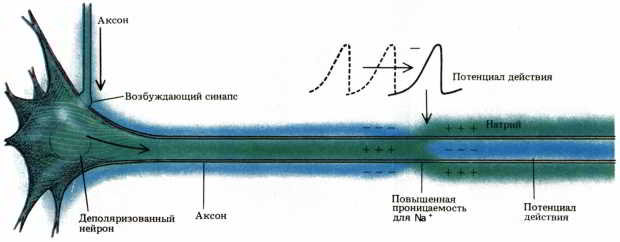
Рис. 30. Когда нейрон активируется приходящим к нему возбуждающим импульсом, волна деполяризации временно меняет знак мембранного потенциала. По мере распространения волны деполяризации вдоль аксона последовательные участки аксона тоже претерпевают эту временную реверсию. Потенциал действия можно описать как поток положительно заряженных ионов натрия (Na+), переходящих через мембрану внутрь нейрона.
Деполяризация, связанная с потенциалом действия, распространяется вдоль аксона как волна активности (рис. 30). Движение ионов, возникающее около деполяризованного участка, способствует деполяризации следующего участка, и в результате каждая волна возбуждения быстро достигает всех синаптических окончаний аксона. Главное преимущество электрического проведения импульса по аксону состоит в том, что возбуждение быстро распространяется на большие расстояния без какого-либо ослабления сигнала.
Кстати, нейроны с короткими аксонами, по-видимому, не всегда генерируют нервные импульсы. Это обстоятельство, если оно будет твердо установлено, может иметь далеко идущие последствия. Если клетки с короткими аксонами способны изменять уровень активности, не генерируя потенциалы действия, то исследователи, пытающиеся по электрическим разрядам оценить роль отдельных нейронов в определенных видах поведения, легко могут упустить из виду многие из важных функций «молчащих» клеток.
Синаптические медиаторы
С некоторыми оговорками синапсы можно сравнить с перекрестками на проводящих путях мозга. В синапсах сигналы передаются только в одном направлении — с концевой веточки посылающего их пресинаптического нейрона на ближайший участок постсинаптического нейрона. Однако быстрая электрическая передача, так хорошо действующая в аксоне, в синапсе не работает. Не вдаваясь в биологические причины этого, мы можем просто констатировать, что химическая связь в синапсах обеспечивает более тонкую регуляцию свойств мембраны постсинаптической клетки.
При общении друг с другом люди передают основное содержание своей речи словами. Чтобы сделать более тонкие акценты или подчеркнуть дополнительный смысл слов, они пользуются тембром голоса, мимикой, жестами. При коммуникации нервных клеток основные единицы информации передаются специфическими химическими посредниками — синаптическими медиаторами (определенный нейрон использует один и тот же медиатор во всех своих синапсах). Если продолжить нашу аналогию с вербальным и невербальным способом общения, то можно сказать, что одни химические посредники передают «факты», а другие — дополнительные смысловые оттенки или акценты.
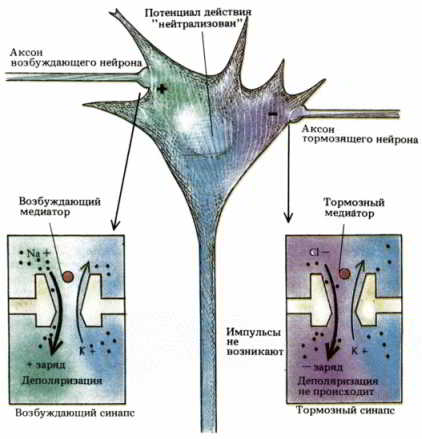
Рис. 31. Противоположное действие возбуждающего (слева) и тормозного (справа) медиаторов можно объяснить тем, что они влияют на разные ионные каналы.
Вообще говоря, существуют два вида синапсов — возбуждающие и тормозные (рис. 31). В первом случае одна клетка приказывает другой переходить к активности, а во втором, наоборот, затрудняет активацию клетки, которой передается сигнал. Под действием постоянных тормозящих команд некоторые нервные клетки хранят молчание до тех пор, пока возбуждающие сигналы не заставят их активироваться. Например, нервные клетки спинного мозга, которые приказывают вашим мышцам действовать, когда вы ходите или танцуете, обычно «молчат», пока не получат возбуждающих импульсов от клеток моторной коры. Под действием спонтанных возбуждающих команд другие нервные клетки переходят к активности, не дожидаясь осознанных сигналов; например, нейроны, управляющие движениями грудной клетки и диафрагмы при дыхании, подчиняются клеткам более высокого уровня, которые реагируют только на концентрацию О2 и СО2 в крови.
Судя по тому, что сегодня известно науке, межнейронные взаимодействия, происходящие в мозгу, можно в основном объяснить, исходя из возбуждающих и тормозных синаптических воздействий. Однако существуют и более сложные модифицирующие воздействия, которые имеют большое значение, так как они усиливают или уменьшают интенсивность ответной реакции нейрона на входные сигналы от различных других нейронов.
Рассмотрим модифицирующие медиаторные сигналы, представив себе, что они носят условный характер. Под термином «условный» подразумевается, что клетки реагируют на них только при определенных условиях, т.е. тогда, когда эти сигналы действуют в сочетании с другими возбуждающими или тормозными сигналами, приходящими по другим путям. Музыканты, например, могли бы считать условным действие педалей у фортепьяно — в том смысле, что для достижения какого-нибудь эффекта их нажатие должно сочетаться с другим действием. Просто нажимать на педали, не ударяя при этом по клавишам, бессмысленно. Звучание ноты изменяется лишь тогда, когда мы нажимаем одновременно и на педаль, и на клавишу. Многие нейронные сети, выполняющие условные функции, — это те, медиаторы которых играют важную роль в лечении депрессии, шизофрении и некоторых других психических расстройств (более подробно эти проблемы обсуждаются в гл. 9).
В заключение несколько слов о процессах, лежащих в основе различных изменений, производимых медиаторами в клетках, на которые они воздействуют. Эти изменения обусловлены ионными механизмами, связанными с электрической и химической регуляцией свойств мембраны. Возбудимость нейрона изменяется потому, что медиатор изменяет поток ионов, переходящих внутрь клетки или же из клетки наружу. Для того чтобы ионы могли проходить через мембрану, в ней должны быть отверстия. Это не просто дыры, а специальные крупные трубчатые белки, называемые «каналами». Некоторые из этих каналов специфичны для определенного иона — натрия, калия или кальция, например; другие не столь избирательны. Некоторые каналы могут открываться с помощью электрических команд (таких, как деполяризация мембраны при потенциале действия); другие открываются и закрываются под действием химических посредников.
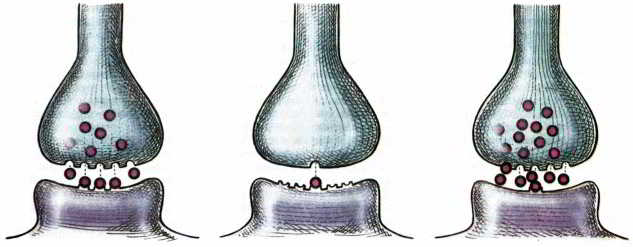
Рис. 32. Схема адаптивных регуляторных процессов, используемых для поддержания нормальной синаптической передачи несмотря на изменения, вызываемые различными препаратами, а также, возможно, заболеваниями. Регулируется количество высвобождаемого или воспринимаемого медиатора. Слева — нормальное состояние. В середине — из-за недостаточного синтеза или сохранения медиатора постсинаптическая клетка увеличивает число рецепторов. Справа — при повышенном выбросе медиатора постсинаптическая клетка уменьшает число или эффективность рецепторов.
Полагают, что каждый химический посредник оказывает на клетки воздействие с помощью химически обусловленных изменений в ионной проницаемости. Определенные ионы и молекулы, используемые тем или иным медиатором, становятся поэтому химическим эквивалентом передаваемого сигнала.
Изменчивость нейронных функций
Как мы видели, нейрон должен успешно справляться с определенными задачами, чтобы функционировать как часть специфической нейронной сети. Медиатор, который он использует, должен передавать определенную информацию. У нейрона должны быть поверхностные рецепторы, с помощью которых он мог бы связывать медиатор в своих входных синапсах. Он должен иметь необходимые запасы энергии для «откачивания» избыточных ионов обратно через мембрану. Нейроны с длинными ветвящимися аксонами должны также транспортировать ферменты, медиаторы и другие молекулы из центральных участков цитоплазмы, где происходит их синтез, в отдаленные части дендритов и аксонов, где эти молекулы будут нужны. Как правило, скорость, с которой нейрон выполняет эти функции, зависит от массы его дендритной и аксонной систем и от общего уровня активности клетки.
Общая выработка энергии — метаболическая активность клетки — может изменяться в соответствии с требованиями межнейронных взаимодействий (рис. 32). Нейрон может повышать свою способность к синтезу и транспортировке специфических молекул в периоды усиленной деятельности. Точно так же при малой функциональной нагрузке нейрон может снизить уровень активности. Эта способность к регуляции фундаментальных внутриклеточных процессов позволяет нейрону гибко приспосабливаться к самым различным уровням активности.
Генетическая детерминация основных типов нейронных сетей
Для того чтобы мозг нормально функционировал, потоки нервных сигналов должны находить надлежащие маршруты среди клеток различных функциональных систем и межрегиональных объединений. В главе 1 мы получили некоторые элементарные сведения о сложном процессе построения и развития мозга. Однако до сих пор остается загадкой, каким образом аксоны и дендрита той или иной нервной клетки растут именно в том направлении, чтобы создавались специфические связи, необходимые для ее функционирования. Между тем тот факт, что конкретные молекулярные механизмы, лежащие в основе многих процессов онтогенеза, еще не раскрыты, не должен заслонять от нас другого, еще более поразительного факта-того, что из поколения в поколение в мозгу развивающихся животных действительно устанавливаются нужные связи. Исследования в области сравнительной нейроанатомии говорят о том, что по фундаментальному плану строения мозг очень мало изменился в процессе эволюции. Нейроны специализированного зрительного рецепторного органа — сетчатки — всегда соединяются с вторичными нейронами зрительной, а не слуховой или осязательной системы. В то же время первичные слуховые нейроны из специализированного органа слуха — улитки — всегда идут к вторичным нейронам слуховой системы, а не зрительной или обонятельной. Точно такая же специфичность связей характерна для любой системы мозга.
Высокая специфичность структуры мозга имеет важное значение. Общий диапазон связей для большинства нервных клеток, по-видимому, предопределен заранее, причем эта предопределенность касается тех клеточных свойств, которые ученые считают генетически контролируемыми. Набор генов, предназначенных для проявления в развивающейся нервной клетке, каким-то еще до конца не установленным образом определяет как будущий тип каждой нервной клетки, так и принадлежность ее к той или иной сети. Концепция генетической детерминированности приложима и ко всем остальным особенностям данного нейрона -например к используемому им медиатору, к размерам и форме клетки. Как внутриклеточные процессы, так и межнейронные взаимодействия определяются генетической специализацией клетки.
Три генетически детерминированных типа нервных сетей
Чтобы сделать концепцию генетической детерминации нейронных сетей более понятной, давайте уменьшим их число и представим себе, что наша нервная система состоит всего лишь из 9 клеток (см. рис. 33). Это абсурдное упрощение поможет нам увидеть сети трех основных типов, которые встречаются повсюду, — иерархические, локальные и дивергентные с одним входом. Хотя число элементов в сетях может быть различным, выделенные три типа могут служить основой для построения надежной классификационной схемы.
Иерархические сети. Наиболее распространенный тип межнейронных связей можно увидеть в главных сенсорных и двигательных путях. В сенсорных системах иерархическая организация носит восходящий характер; в нее включаются различные клеточные уровни, по которым информация поступает в высшие центры — от первичных рецепторов к вторичным вставочным нейронам, затем к третичным и т.д. Двигательные системы организованы по принципу нисходящей иерархии, где команды «спускаются» от нервной системы к мышцам: клетки, расположенные, фигурально говоря, «наверху», передают информацию специфическим моторным клеткам спинного мозга, а те в свою очередь — определенным группам мышечных клеток.
Иерархические системы обеспечивают очень точную передачу информации. В результате конвергенции (от лат converge — сходиться к одному центру) — когда несколько нейронов одного уровня контактируют с меньшим числом нейронов следующего уровня, или дивергенции (от лат. divergo — отклоняюсь, отхожу) — когда контакты устанавливаются с большим числом клеток следующего уровня, информация фильтруется и происходит усиление сигналов. Но, подобно любой цепи, иерархическая система не может быть сильнее своего самого слабого звена. Любая инактивация (от лат. in- — приставка, означающая отрицание) любого уровня, вызванная ранением, заболеванием, инсультом или опухолью, может вывести из строя всю систему. Конвергенция и дивергенция, однако, оставляют цепям некоторый шанс уцелеть даже при их серьезном повреждении, Если нейроны одного уровня будут частично уничтожены, сохранившиеся клетки смогут все-таки поддерживать функционирование сети.
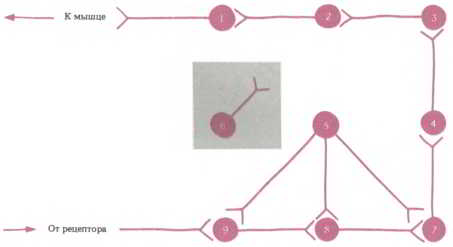
Рис. 33. Нервная сеть из 9 клеток (схема). По периметру — нейроны соединены друг с другом в иерархическую цепь, типичную для сетей сенсорной и двигательной систем. В центре — дивергентная сеть с одним входом (клетки 5, 7, 8, 9), типичная для моноаминэргических систем, в которых один нейрон соединяется с большим числом мишеней. Слева — нейрон локальной сети (6), устанавливающий связи главным образом с клетками из своего ближайшего окружения.
Иерархические системы существуют, конечно, не только в сенсорных или двигательных путях. Тот же тип связей характерен для всех сетей, выполняющих какую-то специфическую функцию, т.е. для систем, которые мы назвали «альянсами» (гл. 1) и более подробно рассмотрим в последующих главах.
Локальные сети. Мы уже говорили о нейронах с короткими аксонами. Если у клетки короткий аксон, настолько короткий, что волнам электрической активности, можно сказать, некуда распространяться, очевидно, что задачи и сфера влияния такого нейрона должны быть весьма ограниченными. Нейроны локальных сетей действуют как фильтры, удерживая поток информации в пределах какого-то одного иерархического уровня. Они, по всей видимости, широко распространены во всех мозговых сетях.
Локальные сети могут оказывать на нейроны-мишени возбуждающее или тормозящее действие. Сочетание этих особенностей с дивергентным или ковергентным типом передачи на данном иерархическом уровне может еще более расширять, сужать или снова фокусировать поток информации.
Дивергентные сети с одним входом. В некоторых нервных сетях имеются скопления или слои нейронов, в которых один нейрон образует выходные связи с очень большим числом других клеток (в таких сетях дивергенция доведена до крайних пределов). Изучение сетей такого типа начато лишь недавно, и единственные места, где они встречаются (насколько нам сейчас известно), — это некоторые части среднего мозга и ствола мозга. Преимущества подобной системы в том, что она может оказывать влияние на множество нейронов сразу и иногда осуществлять связь со всеми иерархическими уровнями, нередко выходя за пределы специфических сенсорных, двигательных и других функциональных альянсов.
Поскольку сфера воздействия таких сетей не ограничена какой-либо системой с определенными функциями, дивергирующие пути этих сетей иногда называют неспецифическими. Однако ввиду того, что такие сети могут влиять на самые различные уровни и функции, они играют большую роль в интеграции многих видов деятельности нервной системы (см. гл. 4). Иными словами, такие системы выступают в роли организаторов и режиссеров массовых мероприятии, руководящих согласованными действиями больших групп людей. Кроме того, медиаторы, используемые в дивергентных системах с одним входом, — это медиаторы с «условным» действием: их эффект зависит от условии, в которых он осуществляется. Подобные воздействия весьма важны и для интегративных механизмов (лат. integratio — восстановление, восполнение, от integer — целый). Однако дивергентные сети такого типа составляют лишь небольшую часть всех нервных сетей.
Изменчивость генетически детерминированных типов сетей
Хотя общая картина связей специфических функциональных сетей удивительно сходна у всех, представителей одного вида, опыт каждой отдельной особи может оказывать дальнейшее влияние на межнейронные связи, вызывая в них индивидуальные изменения и корректируя их функцию.
Представим себе, например, что в мозгу большинства крыс каждый нейрон 3-го уровня в зрительной системе соединен примерно с 50 клетками-мишенями 4-го уровня — сравнительно небольшая дивергенция в системе, характеризующейся в остальном четкой иерархией. Теперь посмотрим, что произойдет, если крыса вырастет в полной темноте? Дефицит входной информации приведет к перестройке зрительной иерархии, так что каждый нейрон 3-го уровня будет контактировать только с 5 или 10 нейронами 4-го уровня вместо обычных 50. Однако, если мы рассмотрим нейроны 4-го уровня в микроскоп, мы убедимся, что у них нет недостатка во входных синапсах. Хотя зрительные нейроны 3-го уровня из-за малого числа связей передают информацию на 4-й уровень в ограниченном объеме, ее дефицит восполняется за счет других работающих сенсорных систем. У нашей крысы в доступном синаптическом пространстве 4-го уровня происходит процесс расширенной переработки слуховой и обонятельной информации.
Рассмотрим другой случай, где тот же эффект проявляется не столь резко. По некоторым данным, интенсивность межнейронной передачи сигналов может влиять на степень развития синаптических контактов между уровнями. Ряд ученых придерживается мнения, что некоторые формы памяти обусловлены изменениями в эффективности таких контактов. Эти изменения могут быть связаны как с микроструктурой (увеличение или уменьшение числа синапсов между клеткой А и клеткой Б), так и с действием медиаторов, участвующих в передаче сигналов (изменение количеств медиатора, синтезируемых и высвобождаемых одной клеткой, или степени реактивности другой клетки) (см. выше рис. 32). Эта тонкая регулировка локальных синаптических функций очень важна при некоторых заболеваниях мозга, о природе которых нам мало что известно (см. гл. 9). Малейшие изменения, происходящие на уровне синаптической активности, могли бы действительно вызвать аномалии поведения, но изменения эти настолько малы, что трудно установить, какова их роль на самом деле.
Нервные клетки не уникальны в своей способности к функциональным изменениям. Во многих других тканях клетки тоже могут изменяться, приспосабливаясь к нагрузке. Если мы возьмем небольшую пробу ткани из четырехглавой мышцы бедра у начинающего тяжелоатлета, а затем у него же после нескольких месяцев усиленной тренировки, то увидим, что каждое мышечное волокно содержит теперь сократимые фибриллы несколько большего размера и число этих фибрилл увеличилось. Слущивающиеся старые клетки вашей кожи и те, что выстилают желудочно-кишечный тракт, ежедневно заменяются новыми; эти клетки, однако, обладают способностью, которой нет у нейронов — они могут делиться. Нейроны генетически запрограммированы на синтез специфических молекул, с помощью которых работают синапсы, а также на образование весьма специфических связей, но не способны к делению. Представьте, что было бы, если бы нервные клетки стали делиться после образования синаптических связей. Как смогла бы при этом клетка распределить свои входные и выходные сигналы, чтобы сохранить прежние связи?
Хотя нейроны и не могут делиться, они обладают большей по сравнению с другими клетками способностью к адаптивной перестройке. Как показали эксперименты, в которых удаляли небольшой участок мозга, а затем в течение нескольких недель наблюдали за реакцией оставшихся частей, некоторые нервные клетки действительно могут регулировать степень своей связи с мишенями. Как правило, при повреждении некоторых синапсов одного нейрона другие, неповрежденные нейроны могут восполнить утраченные звенья цепи путем некоторого ускорения нормального процесса замены синапсов. Если две нервные клетки должны «общаться» более интенсивно, число связей между ними может возрастать за счет добавления новых синапсов при одновременном сохранении старых.
По-видимому, статичность макроскопического строения нервной системы заслонила от нас факт постоянного роста и отмирания связей. Существует даже мнение, что нейроны в нормальном состоянии все время образуют новые связи со своими мишенями. Как только новые синапсы сформировались, старые разрушаются. Такое замещение, вероятно, может компенсировать изнашивание связей в результате их длительной и непрерывной работы.
Хотя испытанное временем представление о том, что наш мозг не может регенерировать утраченные клетки, остается по-прежнему справедливым, исследования последних лет наводят на мысль, что здоровые нейроны обладают значительной структурной пластичностью. Этот более динамичный взгляд на изменчивость мозга открывает широкое поле для исследований; но прежде чем мы начнем понимать, как могут меняться синаптические связи, нам предстоит еще многое узнать.






