Глава 1. Анатомо-физиологические особенности поджелудочной железы у детей
Анатомо-физиологические особенности поджелудочной железы у детей чаще всего обусловлены незаконченным развитием ее к моменту рождения. Многие заболевания этого органа протекают у детей не так, как у взрослых, что можно объяснить не только общими свойствами детского
организма, но и возрастными особенностями строения поджелудочной железы и ее функции. Врожденная патология, чаще наблюдаемая у детей, связана с нарушением нормального развития pancreas.
Сведения об эмбриональном и постнатальном развитии поджелудочной железы у человека начали появляться после первой обобщающей работы Helly (1900). В дальнейшем в исследованиях О. Гертвига (1908), Corning (1922), Nakamura (1924), Neubert (1927) приводятся почти идентичные данные по этому вопросу, совпадающие с современными представлениями (Н. И. Лепорский, 1951; А. Г. Кнорре, 1967; М. Л. Ногаллер, 1968; Patten, 1946; Seifert, 1956).
По данным большинства авторов, в первый месяц эмбрионального развития у человеческого эмбриона 3—5 мм длины появляются закладки поджелудочной железы в виде эндодермальных выпячиваний верхнего отдела средней части первичной кишечной трубки. Различают большой непарный дорсальный и вентральные зачатки (правый и левый). Вентральный левый зачаток вскоре исчезает, в то время как правый, перемещаясь налево и кзади, сливается с дорсальным зачатком на 2-м месяце эмбрионального развития у зародыша длиной 15 мм (рис. 1).
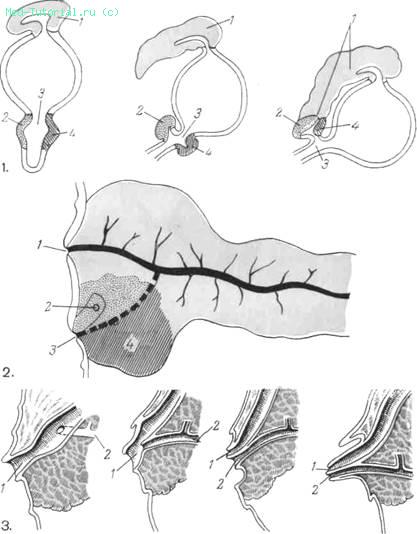
1. Схема развития поджелудочной железы из вентральных и дорсального зачатков (из Corning). I — дорсальный зачаток; 2, 4 — задний и передний вентральные зачатки; 3 — желчный проток. Пояснения в тексте.
2. Зоны формирования поджелудочной железы из вентрального (заштрихованы: 4 — задний, точками — передний) и дорсального зачатков. 1 — добавочный проток (Santorini); 2— желчный проток; 3 — панкреатический проток (Wirsungi).
3. Варианты впадения панкреатического и желчного протоков (по Letulle,1898).1 — желчный проток; 2— панкреатический проток.
При наливках проводящих протоков в этом эмбриональном периоде (Lecco, 1910) было выявлено, что хвост, тело и проксимальная часть головки с d. pancreaticus minor (Santorini) происходят из дорсальной закладки, а остальная часть головки и d. pancreaticus major (Wirsungi)—из вентральной (рис. 2). В дальнейшем d. Santorini чаще всего исчезает и остается d. Wirsungi.
Сложное развитие поджелудочной железы обусловливает множественные варианты нормального строения протоковой системы и особенно места впадения выводного протока в двенадцатиперстную кишку. Вариабельность впадения панкреатического протока в области дуоденального ссека многими рассматривается как норма во всех случаях (рис. 3). Возникшая было теория «общего канала», считающая строение p. Vateri в виде единой ампулы для d. choledochus и d. pancreaticus причиной возникновения панкреатитов (заброс желчи в панкреатический проток), в последнее время вызывает ряд возражений.
Определенный интерес представляет развитие сложной гистологической структуры поджелудочной железы. В начале
3-го эмбрионального месяца возникают только толстостенные протоки с боковыми ветвями без признаков внешней секреции и без дифференцирования ацинусов и островков. С конца 3-го месяца эмбрионального периода тело железы расчленяется на различные железистые дольки с протоками, выстланными цилиндрическим эпителием, появляются зародышевые почки, расположенные кнутри и кнаружи от протоков, а также продуцирующие секрет клетки. Из этого так называемого аденомера развиваются ацинусы, островковая ткань и протоковая система. Нарастание массы железы происходит в зависимости от длины и массы плода и новорожденного (табл. 1), а также возраста ребенка (рис. 4).
Таблица 1 Масса поджелудочной железы у новорожденных (Potter, 1953)
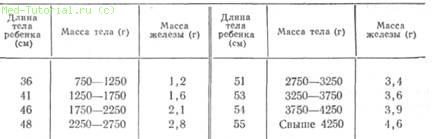
Инсулярный аппарат в начале процесса развития составляет большую часть массы pancreas. У эмбриона 20 см длиной масса инсулярного аппарата и мезенхимы составляет 2/з массы железы, т. е. в это время поджелудочная железа представляет собой железу внутренней секреции. После рождения инкреторная часть составляет уже 30%, уменьшаясь в процессе дальнейшего роста до 3—4%, как у взрослых людей.
Экзокринный аппарат, развиваясь, постепенно вытесняет соединительную ткань, составляющую значительно больший процент массы железы у детей после рождения. Только к 2 годам гистологическое строение поджелудочной железы начинает приближаться к таковому у взрослых.
Таким образом, к особенностям развития поджелудочной железы у детей можно отнести относительную незрелость экзокринной части после рождения и мощное раннее развитие функционирующего эндокринного аппарата.
По гистологическому строению поджелудочная железа относится к сложным трубчатоальвеолярным железам. Она состоит из железистой и соединительной ткани.
Железистая ткань представлена дольками размером до 5 мм (А. В. Смирнов, О. Б. Порембский, Д. И. Фрид, 1972), внутри которых находятся клетки, вырабатывающие панкреатический сок, и особые скопления клеток в виде островков Лангерганса, несущих эндокринную функцию. Долька окружена соединительной тканью, где расположены кровеносные и лимфатические сосуды и нервы.
Экзокринная часть поджелудочной железы на всём своем протяжении от выводного протока до концевых отделов — однослойна. Это Связано с тем, что железа происходит из типичного однослойного эпителия слизистой оболочки кишки.
Концевые отделы секреторной части имеют альвеолярное или алввеолярно-трубчатое строение.
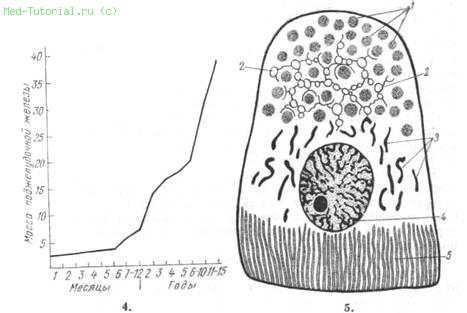
Клетки концевых отделов производят большое количество белковых веществ с разным строением белковых молекул. Их строение находится в тесной связи с функцией. Клеточное ядро, играющее роль регулятора и контролера обмена веществ в клетке, во время клеточного синтеза белка пересылает в цитоплазму нуклеолярную субстанцию и при этом циклично меняется. Цитоплазма ацинозных клеток имеет следующее гистологическое строение: между ядром и основанием клетки находится эргастоплазма с базальными нитями (хондриозомами), эргастоплазма базофильно окрашивается, состоит из рибонуклеиновых кислот и содержит щелочную фосфатазу; между ядром и апикальной частью клетки находятся эозинофильно окрашивающиеся гранулы проферментов («зимогенные гранулы») и аппарат Гольджи; в окружности ядра расположены митохондрии, в которых локализуются дыхательные ферменты и ферменты цикла Кребса (рис. 5).
4. Нарастание веса железы в зависимости от возраста.
5. Строение ацинарной клетки (по Seifert). / — ядрышки проэнзима; 2 — аппарат Гольджи; 3 — митохондрии; 4 — ядро с ядрышком; 5 — эргастоплазма (базальные нити).
В выделительную фазу нуклеолярная субстанция (белок и рибонуклеиновая кислота) транспортируется по хромосомным проводящим каналам в цитоплазму (при этом клеточное содержимое и цитоплазма не смешиваются). Белок и рибонуклеиновая кислота используются в эргастоплазме (может быть также в митохондриях) для пополнения нуклеопротеида, входящего в состав цитоплазматического секрета. Последующая фаза восстановления характеризуется закрытием мембранных пор ядра, исчезновением каналов, центрипетальным транспортом оставшейся нуклеолярной субстанции, уменьшением объема ядра и наступающим утолщением видимых ядерных структур за счет увеличения количества дезоксирибонуклеиновой кислоты.
Механизм секреции протекает по двум принципам: разделения функций (в различных областях дольки железы молено встретить фазу накопления и фазу секреции) и ритмичности — клетки одного ацинуса действуют в одинаковом ритме.
Ко времени истощения зимогенных гранул происходит расширение базальной части клетки и утолщение базальных нитей, которые служат местом нахождения резервных веществ для синтеза секрета. Во время синтеза, происходящего при участии ядра, базальная часть клетки уменьшается и базальные нити практически исчезают.
Выделяющийся из клетки секрет попадает в межклеточные секреторные капилляры, а затем в так называемые вставочные отделы, имеющие значительную протяженность. При изучении кровообращения в поджелудочной железе методом наливок тушью Doerr (1952) доказал, что капилляры образуют сеть анастомозов в окружности вставочных отделов и мелких протоков, что подтверждает особую важность этих участков железы. Именно здесь происходит основной обмен между плазмой крови, лимфой (лимфатические сосуды сопровождают кровеносные капилляры) и секретом наподобие функции канальцевой системы нефрона.
Вставочные отделы дольки поджелудочной железы плавно переходят в выводные протоки, выстланные более высоким призматическим эпителием. В стенке выводных протоков находятся слизистые железы.
При изучении возрастной гистологии поджелудочной железы можно отметить значительное преобладание соединительной ткани в дольках поджелудочной железы и между ними у детей грудного возраста и постепенное почти полное ее исчезновение у взрослых (О. Д. Китайгородская, 1931, Emery, 1952).
Вопрос о капсуле поджелудочной железы долгое время дискутировался в иностранной и отечественной литературе (И. М. Топчибашев, 1963; А. В. Смирнов и др.; David, 1955).
В настоящее время общепринятой является точка зрения А. В. Смирнова, О. Б. Порембского и Д. И. Фрида, которые считают, что поджелудочная железа имеет тонкую соединительнотканную капсулу, отдающую внутрь массы железы множественные тонкие отростки. При таком строении можно сказать, что каждая долька железы имеет собственную соединительнотканную капсулу. Если исходить из этих данных, то вряд ли имеет особый смысл рассечение капсулы по передней поверхности железы, рекомендовавшееся раньше для снятия напряжения при отеке pancreas.
Анатомические особенности поджелудочной железы у детей имеют в своей основе как относительную незрелость детского организма и незаконченное развитие органов к рождению ребенка, так и постоянно меняющееся в зависимости от возраста взаимоотношение различных частей тела и органов.
Поджелудочная железа помещается забрюшинно и огибает тела позвонков спереди, располагаясь между fades pancreatica селезенки слева и нисходящей частью двенадцатиперстной кишки. У взрослых различают ложкообразную, молоткообразную и Г-образную форму железы (А. В. Смирнов и др.). В детском возрасте форма поджелудочной железы четко не дифференцируется, но у детей одного и того же возраста уже можно различать либо изогнутую, либо вытянутую форму. На поперечном разрезе железа обычно призматическая или овальная. Располагаясь на проекции позвоночника, pancreas у новорожденных находится на уровне 77г10 и лишь к 7—9 годам спускается к телу I поясничного позвонка, как у взрослых.
Если форма поджелудочной железы на поперечном разрезе призматическая, различают переднюю, заднюю и нижнюю поверхности. Передняя и нижняя поверхность, сливаясь, образуют margo anterior, по которому прикрепляется брыжейка поперечно-ободочной кишки. Длина поджелудочной железы, быстро нарастая с возрастом, довольно вариабельна (табл. 2).
Таблица 2 Колебания длины (в см) поджелудочной железы в различных возрастных группах (по Г. А. Кайсарьянц, 1947)

Головка, тело и хвост поджелудочной железы у детей не всегда отчетливо различаются, однако для удобства изложения и учитывая характерные анатомические особенности, свойственные каждому отделу, мы придерживаемся того же деления, что и у взрослых.
Головка поджелудочной железы расположена в подкове двенадцатиперстной кишки и связана тесно с pars descendens duodeni, в средней трети которой панкреатический проток впадает в p. Vateri, сливаясь с желчным протоком. Взаимоотношения этих двух протоков в месте впадения в двенадцатиперстную кишку описаны выше (см. стр. 5).
Подкова двенадцатиперстной кишки не полностью выполнена головкой pancreas. На границе между pars inferior и pars ascendens duodeni находится отверстие, через которое из корня брыжейки входит v. mesenterica sup., и сливаясь на задней поверхности железы с v. lienalis, образует v. portae. Через это же отверстие выходит a. mesenterica sup., также участвующая в образовании корня брыжейки. Спереди на границе между головкой поджелудочной железы и двенадцатиперстной кишкой имеется видимая борозда, в которой располагается анастомоз a. pancreaticoduodenalis inf. (из a. mesent. sup.) с а. рапсгеа-ticoduodenalis sup., от которого отходят артериальные ветки в головке железы и к двенадцатиперстной кишке. Дорсальную поверхность головки pancreas пересекают v. cava inf. и v. renalis dex., а медиальнее головка приходит в соприкосновение с v. portae, которая, образуясь здесь же из слияния v. lienalis и v. mesenterica sup., продолжает свой путь между листками брюшины в составе lig. hepatoduodenale в ворота печени. Ductus choledochus расположен справа от v. portae в борозде между задней поверхностью головки и двенадцатиперстной кишкой и тянется вплоть до средней трети нисходящей ее части, где сливается с горизонтально исходящим из головки поджелудочной железы d. pancreaticus и впадает в просвет кишки. Часто d. choledochus находится целиком внутри паренхимы головки железы.
Тело и хвост pancreas отделены брюшиной задней стенки сальниковой сумки от желудка. Задняя поверхность поджелудочной железы отделена от аорты рыхлой ретропанкреатической клетчаткой, в которой проходит так называемая собственная связка крючковидного отростка, разделяющая ретропанкреатическую клетчатку на два изолированных пространства. Сам крючковидный отросток в раннем возрасте располагается в одной плоскости со всей железой и лишь с 2 лет поворачивается кзади под острым углом к оси тела железы.
У верхнего ее края от аорты отходит чревная артерия. Задняя поверхность pancreas пересекается в вертикальном направлении справа налево аортой, грудным протоком, непарной веной (см. рис. 6, б).
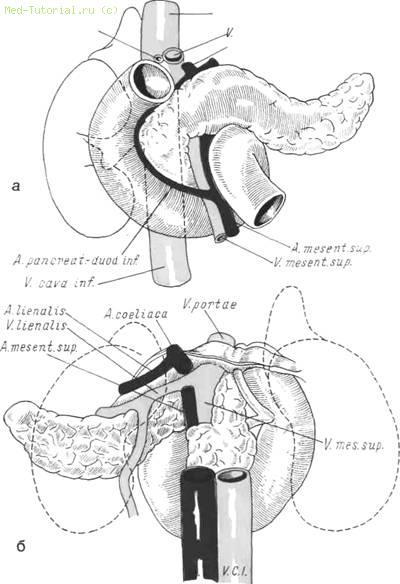
6. Анатомические взаимоотношения поджелудочной железы и двенадцатиперстной кишки. а — вид спереди; б — вид сзади.
Здесь же расположено солнечное сплетение. Селезеночные артерия и вена погружены в неглубокую борозду, проходящую вдоль тела органа. Нижняя поверхность железы узкая и образует край, к которому прикрепляется брыжейка ободочной кишки и прилегает начальный отдел тощей.
Хвост pancreas расположен ретроперитонеально и распространяется вплоть до ворот селезенки, часто плотно с ней соприкасаясь. По верхнему краю хвоста проходят селезеночные артерия и вена, причем борозда, в которой расположены эти сосуды, с возрастом углубляется, и у детей старшего возраста часто трудно резецировать хвост железы, не удалив селезенку. Передняя стенка хвостового отдела принимает участие в образовании сальниковой сумки. У новорожденных задняя поверхность железы в области хвоста пересекает относительно большой надпочечник (с возрастом поле соприкосновения значительно уменьшается). После 7—9 лет хвост pancreas пересекает своей задней поверхностью левую почку.
Связочный аппарат формируется к G—7 годам за счет брюшины, переходящей на соседние органы. Основная масса связок находится между передней поверхностью железы, различными отделами желудка и малого сальника. К этому возрасту появляется и так называемая парапанкреатическая клетчатка, обеспечивающая большую смещаемость железы в области хвоста и части тела.
По данным большинства авторов (А. В. Смирнов и др.; Н. Л. Кущ, А. Д. Тимченко, В. В. Приходченко, 1975), васкуляризацию поджелудочной железы обеспечивают 10 артерий: 5 — поджелудочно-двенадцатиперстных и 5 —поджелудочных (рис. 7).
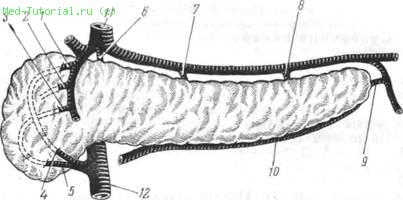
7. Схема артериального кровоснабжения поджелудочной железы.
1 — верхне-задняя поджелудочно-двенадцатиперстная артерия, a. pancreatico-duodena-lis superior posterior; 2 — средняя поджелудочно-двенадцатиперстная артерия, a. pancreaticoduodenalis media; 3 — верхне-передняя поджелудочно-двенадцатиперстная артерия, a. pancreaticoduodenalis superior anterior; 4 — нижнезадняя поджелудочно-двенадцатиперстная артерия, a. pancreaticoduodenalis inferior posterior; 5 — нижнепередняя поджелудочно-двенадцатиперстная артерия, a. pancreaticoduodenalis inferior anterior; 6 — тыльная поджелудочная артерия, a. pancreatica dorsalis; 7 — большая поджелудочная артерия, а. pancreatica magna; 8—пограничная артерия (латинского названия не имеет);. 9 — каудальная поджелудочная артерия, a. pancreatica caudalis;10 — нижняя поджелудочная артерия, a. pancreatica inferior; //—чревная артерия, a. coeliaca; 12—верхне-брыжеечная артерия, a. mesenterica superior.
Первые отходят от ветвей чревной и верхнебрыжеечной артерий, осуществляя кровоснабжение головки pancreas; вторые — от селезеночной, верхнебрыжеечной и желудочно-двенадцатиперстной артерий и питают тело и хвост железы. Необходимо учитывать, что у детей артерии поджелудочной железы отличаются тонкостью и эластичностью. Они образуют мощную сеть анастомозов между артериями tr. coeli-acus и a. mesenterica sup.
Вены поджелудочной железы также делятся на две большие группы: первая группа (поджелудочно-двенадцатиперстные) получает кровь от головки pancreas и двенадцатиперстной кишки; вены второй группы (поджелудочные) несут кровь только от железы. Сосуды впадают в воротную, верхнебрыжеечную и селезеночную вены, сопровождая по ходу одноименные артерии, но располагаясь несколько поверхностнее.
Протоки поджелудочной железы у детей на всем протяжении имеют одинаковый диаметр. Главный выводной проток формируется из слияния внутридольчатых и междольчатых выводных протоков, помещается в толще железы, обычно центрально, но возможны и отклонения от оси органа, чаще кзади. Возрастные изменения длины протока соответствуют изменению размеров самой железы (см. табл. 2). Главный панкреатический проток у места слияния с концевым отделом d. choledochus изгибается вниз и несколько кзади, впадает общим стволом в двенадцатиперстную кишку, открываясь в papilla Vateri (изредка несколько ниже большого дуоденального сосочка). В головке поджелудочной железы располагается добавочный панкреатический проток, который анастомозирует с главным и впадает в двенадцатиперстную кишку выше p. Vateri в области малого сосочка.
Лимфатическая система поджелудочной железы у детей не имеет каких-либо анатомических особенностей. Истоками ее являются тканевые щели, расположенные между клетками по ходу базальной мембраны. Эти межтканевые щелевые
пространства, сливаясь между собой, образуют лимфатические капилляры, которые располагаются на наружной поверхности концевых отделов железы и вместе с капиллярами соседних концевых отделов формируют лимфатические сосуды. Последние, выйдя на поверхность железы, располагаются в междольковых поверхностных бороздах и направляются к регионарным лимфатическим узлам первого этапа.
Для характеристики лимфатических узлов мы считаем наиболее подходящей схему А. В. Смирноеа, О. Б. Порембского и Д. И. Фрида, согласно которой все регионарные узлы поджелудочной железы первого этапа разделены на 8 групп:
Lymphoglandulae lienales — по ходу селезеночных сосудов;
Lymphoglandulae hepaticae — по ходу печеночной артерии;
Lymphoglandulae mesentericae — по ходу верхнебрыжеечной артерии;
Lymphoglandulae pancreaticoduodenalis anteriores — по ходу передней поджелудочно-двенадцатиперстной борозды;
Lymphoglandulae pancreaticoduodenalis posteriores — по ходу задней поджелудочно-двенадцатиперстной борозды;
Lymphoglandulae pancreaticae anteriores — по ходу переднего края поджелудочной железы;
Lymphoglandulae pancreatico-lienales— в области хвоста поджелудочной железы;
лимфоузлы у места слияния общего желчного протока с главным панкреатическим протоком (не имеют латинского названия).
Лимфа из этих узлов может поступать в лимфатическую систему желудка, печени, желчного пузыря, большой сальник, парааортальные узлы и другие органы. Этим объясняется широкое и разнообразное метастазирование при злокачественном поражении поджелудочной железы, а также мощный перипанкреатический воспалительный процесс с включением в инфильтрат многих близлежащих органов при гнойном поражении pancreas.
Иннервация поджелудочной железы осуществляется парасимпатическими и симпатическими волокнами. Первые поступают в составе блуждающих нервов, вторые — чревных нервов. Блуждающий нерв контролирует в основном секреторную деятельность поджелудочной железы. Значительная роль в регуляции ее функции принадлежит ядерным образованиям межуточного мозга, которые обеспечивают взаимосвязь между корой головного мозга и нижележащими структурами. В парасимпатической иннервации участвуют вицеральные ядра блуждающего нерва кранио-бульбарного отдела. Центральный отдел симпатической иннервации образован клеточными группами, расположенными в сером веществе спинного мозга, его боковых рогах. Волокна этих клеток выходят в составе переднего корешка, отделяются от него в виде г. г communicantes albi и входят в узлы пограничного симпатического ствола, из клеток которого начинаются постганглионарные невроны. Симпатические волокна, осуществляющие иннервацию поджелудочной железы, исходят из Di—Dix симпатических узлов, образуя п. splanchnicus major и из D—Dxn узлов — п. splanchnicus minor.
Необходимо учитывать, что спинной мозг до 3-го месяца эмбрионального развития соответствует длине позвоночного канала, но в процессе развития отстает от роста позвоночника и к моменту рождения достигает только Ш поясничного позвонка (у взрослых — границы I——11 поясничных позвонков). Таким образом, сегменты грудного отдела пограничного симпатического ствола располагаются на 2 позвонка выше соответствующего симпатического узла (М. Б. Цукер, 1970; А. В. Триумфов, 1974, и др.)- Большие и малые чревные нервы подходят к солнечному сплетению и оттуда дают ветви ко всем органам брюшной полости. Симпатические волокна (в числе которых имеются чувствительные) правого и левого симпатического ствола раздельно иннервируют головку, тело и хвост pancreas (White, 1966), они контролируют кровоснабжение железы и ингибируют ее секрецию.
Постганглионарные и парасимпатические нервные волокна, идущие от солнечного сплетения, расположены по ходу желудочно-двенадцатиперстной артерии и нижней части общего желчного протока по направлению к головке железы и сфинктеру Одди, а также походу селезеночной артерии — в направлении к телу и хвосту pancreas. Вокруг поджелудочной железы чревные и блуждающие нервы образуют пять нервных сплетений (солнечное, печеночное, верхнебрыжеечное, селезеночное и левое почечное), отдающих нервные волокна в поджелудочную железу (А. А. Шелагуров, 1970; А. В. Смирнов и др.; Scholze, 1972).
Развитие периферической симпатической и парасимпатической нервной системы к моменту рождения ребенка не заканчивается. Для внутриутробного периода характерен рассыпной тип строения солнечного сплетения. У новорожденных и детей до 1 года нервные волокна не только сопровождают сосуды, но и направляются непосредственно в паренхиму самостоятельными стволиками. Появление крупных сплетений можно определить лишь к 2-летнему возрасту, а часто и значительно позднее. Время образования нервных сплетений индивидуально и бывает чрезвычайно замедлено (Р. С. Новоселова, 1940). Это определяет несколько иное отношение к околопочечной новокаиновой блокаде по А. В. Вишневскому у детей младшего возраста, так как отсутствие крупных нервных сплетений у них не всегда позволяет добиться эффекта, подобного получаемому у взрослых, а увеличение количества новокаина, вводимого в забрюшиниую клетчатку, может вызвать и нежелательное токсическое действие.
Физиологические особенности поджелудочной железы — экзокринно-эндокринного органа — начинают проявляться у ребенка внутриутробно и в дальнейшем меняются в процессе роста. Ферменты, образуясь в ацинарных клетках, скапливаются в них в виде гранул, а выход гранул, содержащих секрет, в ацинарный проток происходит после слияния их с мембраной клетки. В дальнейшем осуществляется интенсивный обмен между образующимся соком, кровью и лимфой в области вставочных отделов. Жидкая часть сока секретируется цилиндрическим эпителием протоков. В главном панкреатическом протоке добавляется мукоидная жидкость, выделяющаяся слизистыми клетками. Pancreas выделяет за сутки в двенадцатиперстную кишку такое количество сока, которое превышает в 10——15 раз массу самого органа. Это указывает на большую секреторную активность железы. Характер секрета изменяется в зависимости от состава пищи, рН его колеблется от 7,1 до 8,2. Панкреатический сок состоит из неорганических веществ (Na, К, Са, Mg, P) в виде карбонатов и бикарбонатов, органических — альбуминов, глобулинов и ферментов.
Различают четыре группы ферментов:
I. Протеазы (пептидазы): трипсиноген, химотрипсиноген, карбоксипептидаза, аминопептидаза, коллагеназа, эластаза.
II. Липаза (эстераза).
III. Карбогидразы (гликозидазы): амилаза, мальтаза, лактаза.
IV. Нуклеазы: рибонуклеаза, дезоксирибонуклеаза.
Первые ферменты появляются на 5-м месяце антенатального
развития в виде трипсиногена (Gschwinc, 1950). Werner (1948) установил, что у плодов от 1000 до 2000 г протеазы содержатся в поджелудочной железе только в небольших количествах, а иногда и отсутствуют, в то время, как у доношенных новорожденных всегда можно обнаружить и ферменты. В работах М. С. Маслова (1952), Sauerbrei и Starke (1949) отмечается, что с возрастом увеличивается выделение секрета поджелудочной железы в двенадцатиперстную кишку.
Количество протеаз в железе и дуоденальном содержимом максимально поднимается к двум годам и незначительно снижается к 5-летнему возрасту до норм взрослых.
Липаза отсутствует у недоношенных новорожденных с массой до 2000 г. У доношенных детей в первые дни жизни концентрация липазы в дуоденальном соке низкая, но в дальнейшем быстро повышается и к возрасту одного года становится в 2,5 раза большей, чем у взрослых. Только к 5 годам концентрация липазы дуоденального сока соответствует нормам взрослого.
Наиболее показательным в возрастном аспекте нам кажется колебание в самой железе и в дуоденальном соке концентрации амилазы. У недоношенных новорожденных в 70% случаев амилаза отсутствует. У зрелых новорожденных цифры амилазы в железе длительное время чрезвычайно низки. К возрасту 1 года концентрация амилазы в железе и дуоденальном содержимом достигает лишь половины цифр взрослых и только к 5 годам, увеличиваясь очень постепенно, становится такой же, как у взрослых. Подобное постепенное нарастание содержания амилазы в поджелудочной железе необходимо учитывать при диагностике заболеваний pancreas у детей в возрасте до 5 лет.
После операций в области поджелудочной железы и непосредственно на ней у новорожденных с высокой кишечной непроходимостью мы ни разу не встречались с подъемом амилазы крови и мочи, хотя проведение специфической антиферментной терапии при подозрении на повреждение pancreas во время операции и соответствующей клинике всегда резко улучшало состояние новорожденного.
Сведения по возрастным изменениям ферментной активности поджелудочной железы у детей в литературе немногочисленны и весьма разноречивы (М. С. Маслов, 1948; Г. М. Монтвилайте, И. И. Садаускас, 1972; Sauerbrei, Starke; Gschwind; Emery; Behrendt, 1962; Тодоров, 1968). Трудность обработки литературы связана с разными методами определения активности ферментов и большими колебаниями цифр в зависимости от вскармливания и функционального состояния желудочно-кишечного тракта.
Все ферменты выделяются в просвет двенадцатиперстной кишки в той концентрации, которая необходима для переваривания поступающей пищи. Регуляция панкреатической секреции осуществляется нервным и гуморальным путем.
Раздражение блуждающего нерва вызывает отделение сока в меньшем количестве, бедного основаниями (бикарбонатами), но богатого белком и ферментами. Препараты, блокирующие блуждающий нерв в области постганглионарных волокон, в частности М-холинолитик атропин, обладают ингибирующим действием на функцию поджелудочной железы. При электронномик-роскопическом анализе секреторного цикла железы в условиях воздействия атропина выяснено, что атропин блокирует выделение пищеварительных ферментов экзокринными клетками в течение 5 ч (Н. К. Пермяков, А. Е. Подольский, 1973). При этом атропин не оказывает влияния на поступление в клетку необходимых для синтеза веществ и синтез ферментов в ацинарной клетке, а действие его заключается в задержке транспорта ферментов внутри клетки и выброса готовой продукции.
Многочисленными исследованиями физиологов начала XX века было доказано, что соляная кислота, поступая в двенадцатиперстную и тощую кишку, стимулирует выработку слизистой оболочкой последней гормонов (секретин, панкреозимин, гепа-токринин, энтерокринин, холецистокинин), которые, всасываясь в кровь, активируют панкреатическую секрецию. Секретин, поступая в кровеносное русло, действует на рецепторы кровеносных сосудов, повышая объем секреции pancreas в 6—8 раз, концентрацию бикарбонатов — в 4—5 раз, но концентрация протеолитических ферментов при этом уменьшается в 3—4 раза (т. е. количество выделяющихся ферментов не меняется). При выключении симпатической иннервации каким-либо способом действие секретина блокируется (А. В. Соловьев, 1959). Панкреозимин также увеличивает количество отделяющегося сока в 2 раза, причем содержание в нем белка возрастает в 4 раза, а трипсина — в 6 раз.
При рассмотрении нормальной физиологии поджелудочной железы необходимо учитывать, что понятия о возрастной норме нельзя рассматривать в отрыве от обычных соотношений питательных веществ и от потребности в белках, жирах и углеводах. Питание детей в разные возрастные периоды предъявляет различные требования к ферментативной деятельности поджелудочной железы. Ребенок от 1 года до 4 лет должен получать на каждый килограмм массы 3,5 г белка, 4—4,5 г жиров и 12—15 г углеводов, в то время как взрослым необходимо 1—2 г белка, 1,5 — жиров, 6,5 — углеводов на килограмм массы (А. А. Шелагуров). Из этого следует, что воздействие различных видов диеты — углеводной, белковой, жировой — должно рассматриваться, исходя из повышенных требований детского организма к этим ингредиентам. Диета, которая для взрослого содержит слишком большое количество жиров или белков, может быть нормальной для ребенка. Особенно это касается, как уже говорилось выше, детей в возрасте до 4 лет.
Преимущественно углеводное питание стимулирует выработку секрета, наибольший выброс которого наблюдается при этом через 1,5 ч после приема пищи. При длительном (более 5 дней) углеводном питании могут быть изменения в ультраструктуре ацинуса (Н. К. Пермяков, А. Е. Подольский, Г. П. Титова, 1973), относящиеся к серьезным нарушениям метаболизма клетки.
Преимущественно жировое вскармливание приводит к постепенной утрате клетками своего биологического ритма. Железа как бы работает «на износ». При односторонней жировой диете может наблюдаться даже развитие панкреатита.
При преимущественно белковой диете наблюдается выброс секрета, богатого белковыми веществами и трипсином. Однако накопление секрета в клетке идет параллельно с его выбросом. При длительном белковом вскармливании ультраструктурное поражение клеток ацинуса не наблюдается. Секреция продолжается в увеличенном количестве со своевременным воспроизводством.
Таким образом, можно отметить, что на поздних сроках преимущественно углеводного или жирового питания наблюдаются почти равнозначные дистрофические изменения в поджелудочной железе, возникновение которых связано с дефицитом белка.
Из этого следует, что оптимальным для железы является сбалансированное питание. При этом исключение из рациона различных питательных веществ не должно касаться общего количества белка.
Это согласуется с наблюдениями Wohlgemuth (1907), который также отмечал снижение секреции поджелудочной железы при жировом питании — «диете Вольгемута». Тормозящее действие жира на секрецию можно использовать на практике, не забывая при этом об указанных недостатках преимущественно жировой диеты и обязательно включая в питание белок. Значительным сокогонным действием обладают растворы овощных соков, в то время как цельные, неразведенные соки угнетают секрецию.
Внутренняя секреция поджелудочной железы осуществляется островками Лангерганса и подробно освещена в соответствующей литературе по эндокринологии. Мы остановимся на основных проявлениях внутренней секреции железы, которые необходимо знать для обследования и лечения детей с хирургическими заболеваниями pancreas. С функцией островкового аппарата связано выделение инсулина, глюкагона и липокаина. Выработка калликреина не относится исследователями к деятельности определенных клеточных элементов железы, однако pancreas является одним из основных продуцентов этого важного вещества, действие которого раскрывается нами в главе «Острые панкреатиты».
Выработка инсулина происходит в клетках островков Лангерганса. Физиологическое действие инсулина многогранно. Внешним проявлением его отсутствия является повышение уровня сахара крови. Однако для оценки деятельности островкового аппарата железы необходимо помнить о взаимном тесном влиянии на количество сахара крови функции гипофиза, надпочечников, инсулярного аппарата и печени. Кроме того, содержание сахара непосредственно связано с выделением островковыми клетками железы глюкагона. По мнению А. А. Шелагурова, глюкагон и инсулин являются антагонистами. Глюкагон способствует выделению глюкозы в кровь из запасов гликогена печени. Секреция этих гормонов и взаимодействие регулируются колебаниями содержания сахара в крови.
Липокаин — вещество, препятствующее жировому перерождению печени, по мнению Н. И. Лепорского, выделяется островковым аппаратом железы. Однако введение одного панкреатического сока собакам с удаленной поджелудочной железой также предупреждает развитие жирового перерождения печени, что говорит против эндокринной природы липокаина.
Из особенностей возрастной физиологии островкового аппарата необходимо отметить гипогликемию новорожденных и детей первого года жизни (Т. В. Ломова, 1958; Brock, 1954, и др.). В момент рождения содержание сахара соответствует количеству его у матери (100±20 мг%). После родов содержание сахара крови новорожденного быстро падает и в первые 3—6 ч доходит до 60 мг°/о, в дальнейшем постепенно повышаясь (рис.8).
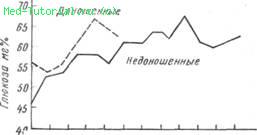
8. Средняя концентрация глюкозы крови у доношенных и недоношенных детей в первые две недели жизни (по И. Тодорову).
После первого года показатели сахара крови колеблются в зависимости от возрастного периода (рис. 9). Между концентрацией инсулина и колебаниями сахара крови нет прямой зависимости (Seifert). Изменения как возрастные, так и патологические зависят в большей степени от экстраинсулярных факторов (Mayer, 1950).
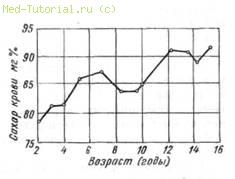
9. Возрастные различия средних нормальных концентраций сахара
крови (по Mayer, 1950).
Следует помнить, что в поджелудочной железе вырабатываются так называемые антиферменты, препятствующие проявлению активного начала фермента. Фермент и антифермент в норме находятся в постоянном равновесии.
