Анатомия и физиология межполушарных различий
В 1968 году в результате детального посмертного исследования мозга у 100 человек Норман Гешвинд и Уолтер Левитски сообщили о заметных анатомических различиях между полушариями. В 65% случаев участок коры височной доли, перекрывающийся с зоной Вернике и называемый planum, был больше в левом полушарии и в 11% случаев — в правом полушарии. В 24% случаев различий между полушариями по этому признаку обнаружено не было. В последующие годы была изучена и измерена не одна сотня других препаратов мозга. Полученные данные в основном совпадают; приблизительно в 70% случаев planum temporale в левом полушарии крупнее (рис. 149). В других исследованиях был обнаружен еще ряд проявлений асимметрии, большая часть которых коррелирует с функциональной право- или леворукостью (табл. 8.3).
Таблица 8.3. Частота (в процентах) анатомических различий между полушариями среди праворуких и леворуких индивидуумов, а также людей, одинаково владеющих обеими руками (амбидекстров). (Corballis М. С. Human Laterality, Academic Press, 1983, p. 72.)
| Вид асимметрии | Праворукие | Леворукие и амбидекстры | ||||
|---|---|---|---|---|---|---|
| да | нет | обратное соотношение | да | нет | обратное соотношение | |
| Сильвиева борозда выше справа (Galaburd, LeMay, Kemper, Geschwind, 1978) | 67 | 25 | 8 | 20 | 70 | 10 |
| Задний рог бокового желудочка длиннее слева (McRae, Branch, Milner, 1968) | 60 | 30 | 10 | 38 | 31 | 31 |
| Лобная доля шире справа (LeMay, 1977) | 61 | 20 | 19 | 40 | 33 | 27 |
| Затылочная доля шире слева (LeMay, 1977) | 66 | 24 | 10 | 36 | 48 | 26 |
| Лобная доля выдается справа (LeMay, 1977) | 66 | 20 | 14 | 35 | 30 | 35 |
| Затылочная доля выдается слева (LeMay, 1977) | 77 | 10,5 | 12,5 | 35 | 30 | 35 |
Подобная асимметрия может быть физической основой функциональных различий между двумя полушариями. Когда впервые появились данные о межполушарных различиях, некоторые ученые высказали предположение, что асимметрия в строении речевых зон может развиваться вследствие обучения языку. Однако такая же асимметрия свойственна и мозгу человеческого плода. Таким образом, анатомическое различие — скорее причина, нежели следствие. Еще одна асимметрия была обнаружена при ангиографии сонных артерий — рентгеновского исследования мозга после введения контрастного вещества в сонную артерию (Galaburda et al., 1978). Известно, что ход крупных сосудов мозга отражает анатомию окружающей ткани. Благодаря такому соответствию удалось установить, что сильвиева борозда (иногда ее называют латеральной бороздой) — глубокая щель в коре мозга, отделяющая височную долю от остальной коры, — в левом полушарии более длинная и более прямая, а в правом она сильнее изогнута вверх. Такая асимметрия выявилась и при изучении ископаемых черепов неандертальского человека; это позволяет предположить, что асимметрия полушарий, вероятно, составляет часть генетического наследия Homo sapiens.
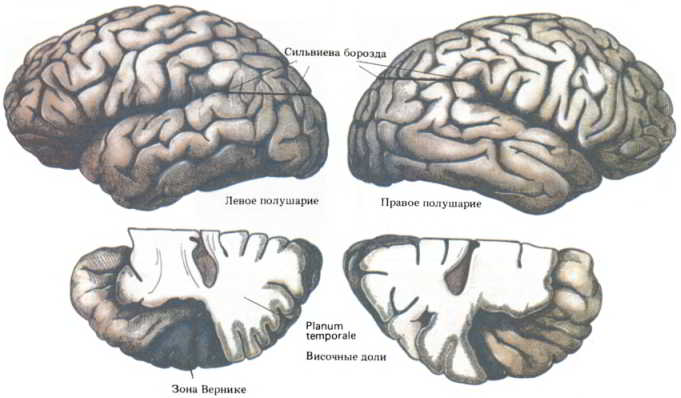
Рис. 149. Анатомическая асимметрия полушарий мозга. Вверху: сильвиева борозда в правом полушарии отклоняется вверх под большим углом. Внизу: задняя часть planum temporale обычно гораздо больше в левом полушарии, связанном с речевыми функциями.
Данные о том, что асимметрия строения мозга у новорожденных детей отражает функциональные различия, получены при изучении вызванных потенциалов, возникающих при звуках человеческой речи у младенцев, которым едва исполнилась неделя. Этот метод позволяет измерять реакцию мозга на раздражители. Например, когда человек видит вспышку света, в зрительной области коры почти одновременно возбуждается большое число нейронов, что ведет к изменению мембранных потенциалов у миллионов нервных клеток. Величина этого вызванного потенциала нередко столь значительна, что ее могут зарегистрировать электроды, приложенные к коже головы. Поскольку электроды регистрируют также и случайную активность мозга, для точного измерения вызванного потенциала требуется неоднократное предъявление стимула, а также обработка полученных данных с помощью компьютера, усредняющего результаты и выдающего кривые наподобие изображенных на рис. 150 и 151.
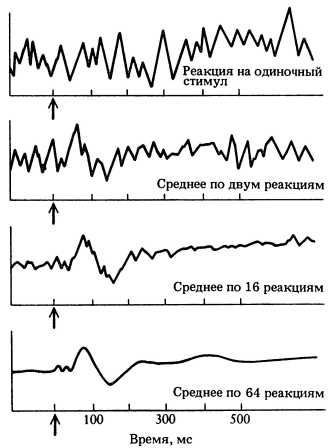
Рис. 150. Вызванный потенциал отражает реакцию мозга на зрительные или слуховые стимулы. Компьютер усредняет отлеты на повторные стимулы, что позволяет выделить вызванную реакцию на фоне обычной электроэнцефалограммы. Усреднение 64 ответных реакций дает четкую картину.
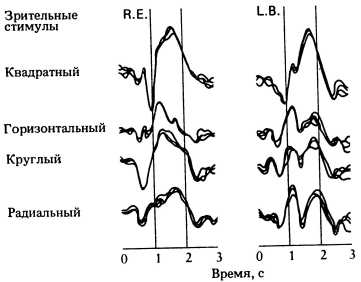
Рис. 151. Реакция индивида на один и тот же раздражитель может быть разной. В каждой колонке представлены ответные реакции одного человека на четыре различных зрительных стимула.
Регистрация электрической активности мозга у младенцев при звуках человеческой речи показала, что у 9 из 10 детей амплитуда реакции в левом полушарии заметно больше, чем в правом. При неречевых звуках — шуме или музыкальных аккордах — у всех 10 младенцев амплитуда вызванных потенциалов была выше в правом полушарии (Molfese et al., 1975).
Таким образом, наш мозг и анатомически, и физиологически, по-видимому, от рождения подготовлен к переработке словесных сигналов. У большинства из нас частью мозга, которая запрограммирована для этой функции, является кора левого полушария. В отличие от большинства сенсорных и двигательных функций процесс становления речевой функции обладает значительной пластичностью. Например, при повреждении речевых областей коры левого полушария в ранний период жизни выполнение их функций берут на себя корковые зоны правого полушария.
В редких случаях, когда, например, все полушарие мозга поражено раковой опухолью, хирургам приходится удалять корковое покрытие целого полушария. Эта процедура носит название гемисферэктомии. У взрослых после удаления правого полушария функция речи почти не нарушается. Те же, кто перенес операцию на левом полушарии, страдают тяжелой формой афазии практически без шансов на улучшение.
Однако если подобная операция производится у маленьких детей, ее результаты бывают совершенно иными. Развитие речи у детей с удаленным в младенческом возрасте левым полушарием идет почти без всяких нарушений. Стандартные тесты по проверке вербальной интеллектуальности не выявляют никаких различий между детьми, перенесшими операцию, и их нормальными сверстниками. Точно так же нет различий между результатами левосторонней и правосторонней гемисферэктомии.
Лишь крайне специализированные тесты все-таки выявляют небольшую разницу. В одном из таких экспериментов трех детей 9-10-летнего возраста, перенесших гемисферэктомию до того, как им исполнилось 5 месяцев, попросили дать оценку трем следующим предложениям с точки зрения смысла и приемлемости:
1) I paid the money by the man[4].
2) I was paid the money to the lady[5].
3) I was paid the money by the boy[6].
У двоих детей было удалено левое полушарие, у третьего — правое. Этот последний с неповрежденным левым полушарием правильно ответил, что первое и второе предложения грамматически неправильны, а третье вполне допустимо. Два его сверстника, перенесшие операцию на левом полушарии, не смогли распознать ошибок, хотя здоровые дети их возраста легко справляются с подобной задачей (Dennis, Whitaker, 1976).
За относительную сохранность способностей у оперированных детей, видимо, ответственна пластичность мозга (см. гл. 7). В ранний период жизни мозг, очевидно, обладает огромными возможностями в отношении собственной перестройки для компенсации ущерба, нанесенного его частям. С возрастом, однако, пластичность уменьшается. Дети более старшего возраста, перенесшие левостороннюю гемисферэктомию, как правило, могут разговаривать, но делают грамматические ошибки и хуже понимают речь.
Пластичность мозга может вступать в действие даже и без повреждения мозга. Когда Джини, девочку, выращенную в изоляции, нашли в возрасте 13 лет (см. гл. 7), она не могла говорить и не понимала, что говорили другие. После того как она до некоторой степени овладела речью, психологи применили метод вызванных потенциалов, чтобы выяснить, какая часть ее мозга ответственна за речь. Они узнали, что как языковыми, так и неязыковыми функциями у Джини управляет правое полушарие.
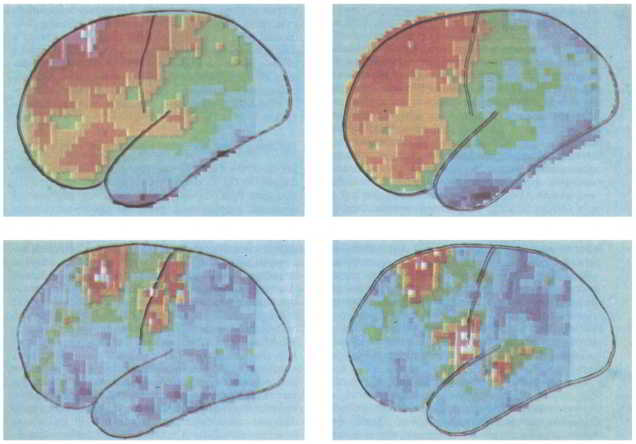
Рис. 152. Один из методов выявления межполушарных различий состоит в измерении кровотока в различных участках мозга при разных видах деятельности. Для этого в кровь вводятся радиоактивные изотопы, и их передвижение регистрируется компьютером, который выдает изображения вроде представленных на этом рисунке. Средней скорости кровотока соответствует зеленый цвет, ниже средней — оттенки синего, выше средней — оттенки красного. Вверху: левое и правое полушария во время покоя при закрытых глазах. Лобные доли обоих полушарий активны, как будто человек думает и что-то планирует. Внизу: левое и правое полушария говорящего человека. Хотя они оба находятся в активном состоянии, зона рта-языка-глотки в правом полушарии менее активна и не отличается в этом отношении от слуховой коры.
Психолингвист Сьюзен Кертис, которая занималась с Джини, полагает, что именно обучение языку играет роль пускового механизма для нормальной специализации полушарий. Если в должное время овладения речью не происходит, «корковая ткань, в норме предназначенная для речи и связанных с нею способностей, может претерпевать функциональную атрофию». Этот вывод основан на результатах исследований Дэвида Хьюбела и Торстена Визеля (см. обсуждение в гл. 3 и 7), показавших, что нейроны, не получавшие нормальных зрительных входных сигналов, не образуют достаточного числа связей с другими клетками и поэтому становятся функционально неактивными. Однако данных, которые указывали бы на неспособность нейронов речевых зон (в случае их бездействия) устанавливать связи, не имеется — эксперименты на животных, подобные опытам Хьюбела и Визеля, не годятся для изучения человеческой речи.






