Возможности и перспективы
Дальнейшая расшифровка физиологической сущности и роли механизма детекции ошибок в мозговой деятельности рассматривается нами как приоритетная задача фундаментальной науки в проблеме «Мозг Человека». Широкое, многоплановое дальнейшее изучение потенциала феномена детекции ошибок («его прав и обязанностей») как общего механизма мозга оказалось возможным, прежде всего, в связи с многочисленностью убедительных свидетельств реальности и надежности проявления феномена. Важно подчеркнуть, что именно приведенные выше свидетельства его реальности позволили изучать влияние этого важнейшего мозгового фактора в условиях, закономерно вызывающих его к жизни, но уже без обязательной одновременной регистрации нейрофизиологического выражения его местных проявлений. Это, как вполне понятно, позволило существенно расширить методические условия работы.
Именно таким образом и было предпринято нами изучение возможного влияния детекции ошибок в вербальном творчестве. Исследование и уточнение роли детектора ошибок в творческом процессе представляет особый интерес, так как по условиям его обнаружения предполагалось, что сигнализация «отхода от известного» должна играть отрицательную, угнетающую роль в деятельности, где важнейшим является фактор новизны (Bechtereva, 1978).
В развитие многолетнего изучения мозговой организации мыслительной деятельности в Институте мозга человека в последние годы (Bechtereva et al., 2004) проведено полиметодическое исследование мозговой организации наиболее сложного ее вида – деятельности творческой. На основе анализа ЭЭГ показано достоверное наличие местных и общих перестроек мозговой активности при реализации соответствующих вербальных психологических тестов. С помощью позитронно-эмиссионной томографии (ПЭТ) в теменно-височной области левого полушария обнаружены зоны, имеющие приоритетное значение для вербального творчества (ПБ 39, 40).
Одновременно обнаружено множество мозговых зон, «обслуживающих» вербальное творчество, то есть описана в основных чертах мозговая структурно-функциональная система организации вербального творчества, что позволило поставить задачу изучения собственно его мозговых механизмов. Исследования, о которых речь пойдет ниже, соединяют две приоритетные линии наших работ – мозгового механизма детекции ошибок и мозговой организации творческой деятельности. В связи с нашими априорными представлениями (см. выше) о том, что активация детекции (детектора) ошибок противостоит творческому процессу, ограничивает выход за рамки (матрицу) уже известного, нами было предпринято психофизиологическое исследование материального (мозгового) выражения этого угнетающего влияния детектора ошибок на вербальное творчество. При такой постановке вопроса мы получали реальную возможность в дополнение к ранее проведенным работам исследовать разные проявленияактивности детектора ошибок, и в частности свойственную некоторым мозговым механизмам и зависящую от конкретной ситуации поливалентность (в данном случае – положительную роль как сигнала об ошибке, отрицательную – как поддерживающего психопатологические проявления при наркоманиях, ограничительную роль в творческом процессе и т. д.).
При дальнейшем изучении детектора ошибок, по-видимому, имеет смысл не забывать о поливалентности многих мозговых механизмов и a priori исключать еще одну его возможную роль в поведенческом контроле и, в частности, в обеспечении нетривиальности и плодотворности творчества. Именно здесь, однако, в экспериментальном решении этой частной задачи, как вполне понятно, могут встретиться значительные методические трудности, прежде всего, в конструировании адекватных психологических тестов.
Исследования мозговых механизмов влияния активации детекции ошибок в вербальном творчестве проводились на группе здоровых испытуемых (25 человек: 14 девушек, 11 мужчин, средний возраст – 23 года). ЭЭГ регистрировалось с поверхности головы испытуемых от 19 отведений, расположенных согласно международной системе 10–20 % в момент выполнения специально разработанных заданий.
Тест состоял из двух пар заданий. В основном тестовом задании (творческое, Тв) испытуемым предъявлялись общеизвестные пословицы и поговорки с отсутствующим последним словом; требовалось вместо пропущенного слова придумать вариант окончания пословицы, возможно более радикально меняющий ее смысл. Контрольное задание (К) в целом было аналогично основному; отличалось требованием вспомнить и назвать недостающее в пословице слово. В тест входили еще два задания, такие же задания, что и в случаях Тв и К, однако тексты пословиц и поговорок в этих заданиях были написаны с орфографическими ошибками (характер ошибок – переставленные буквы или замена одной буквы на другую, т. е. длина слов не изменялась), что предполагало обязательную активацию детектора ошибок (соответственно задания ТвД и КД). Волонтерам требовалось не только выполнить инструкцию к заданиям Тв или К, но и определить количество орфографических ошибок в написании текстов пословиц или поговорок (одна, две, ноль).
В этом исследовании были проанализированы изменения локальной мощности ЭЭГ в различных диапазонах. Статистический анализ данных был осуществлен при использовании дисперсионного анализа ANOVAдиапазон?состояние?зона по плану персональных сравнений (within-subject design). Для иллюстрации различий в отдельных зонах мы использовали post-hoc-анализ с применением LSD критерия Фишера (p<0,05). Достоверные различия были выявлены в ЭЭГ не только для сравнения Тв–К, но и в контрасте ТвД–К. Достоверные различия были представлены в основном в бета-2 (18,5–30 Гц)– и гамма (31–40Гц)-диапазонах ЭЭГ (рис. 4).
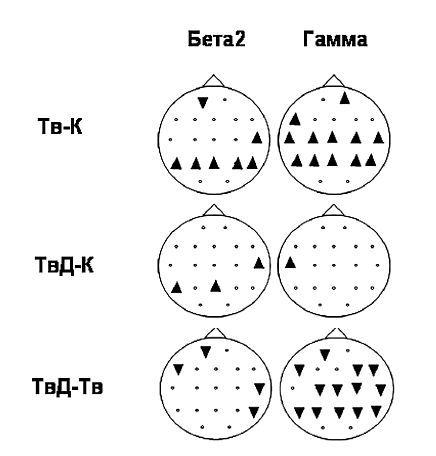
Рис. 4. Изменения мощности ЭЭГ при выполнении творческих заданий «в» и «без» условий внешнего вовлечения в процесс детекции ошибок (p <0,05). Тв – творческое задание; К – задание на воспроизведение из долговременной памяти; ТвД – творческое задание при внешней активации детекции ошибок; КД – задание на воспроизведение из долговременной памяти при внешней активации процесса детекции ошибок треугольник, направленный вверх/вниз, – мощность ЭЭГ была больше/меньше в первом задании из двух сравниваемых
ЭЭГ испытуемых, зарегистрированные в условиях инициации творческой деятельности при корректном предъявлении текстов, характеризовались синхронизацией в гамма-диапазоне практически по всей поверхности коры и синхронизацией в бета2-диапазоне в ассоциативных зонах коры. Активация детектора ошибок (в задании ТвД) привела к исчезновению этого эффекта (сравнения ТвД–К, ТвД–Тв). Корреляты детекции ошибок при сопутствующем решении творческой задачи (сравнение ТвД– Тв) характеризовались локальной десинхронизацией в гаммадиапазоне ЭЭГ в большинстве зон коры. Похожие, но более слабые эффекты были получены в бета-2-диапазоне.
Одно из будущих направлений наших исследований – это разделение эффектов направленного внимания и, собственно, детекции ошибок. Наши начальные результаты выглядят весьма многообещающими, однако они требуют дальнейшего анализа для более детального обсуждения
В заключение следует отметить, что мы ни в коем случае не стараемся умалить важность возможного прогресса в исследованиях негативности, связанной с совершением человеком ошибочных действий (ERN), или других, связанных с событием феноменов в проблеме детекции ошибок. Однако исследование сущности детекции ошибок как общемозгового механизма с применением полиметодического подхода имеет много преимуществ. Мы рассматриваем наши последние результаты как многообещающий шаг в исследованиях роли детекции ошибок в различных видах мыслительной деятельности человека и надеемся на возможность подтверждения гипотезы о детекции ошибок как одном из общемозговых контролирующих механизмов.






