Глава шестая
Нейрофизиология памяти человека и общие механизмы его мозга
И все это может сделать одна только физиология, так как она одна держит в своих руках ключ к истинно научному анализу психических явлений.
Давно и широким фронтом ведется настойчивое многоплановое изучение проблемы памяти. Опыт показал, что достаточно полноценная расшифровка ее механизмов возможна лишь на основе интегративного подхода, но это не исключает отдельных, разных по значимости аналитических этапов ее изучения.
Физиологическое изучение памяти базируется главным образом на исследовании тех изменений различных физиологических показателей в мозгу, которые происходят в ходе запоминания (обучения), динамики следовых явлений и отражения процесса считывания из долгосрочной памяти. Ценные данные о механизмах памяти у человека были получены, как известно, при клинико-анатомических сопоставлениях (Бехтерев, 1900, и многие другие), в процессе развития нейропсихологии (Лурия, 1977; Хомская, 1977) и при анализе результатов электрических воздействий на кору и различные глубокие образования мозга (Ojemann, 1978a; Смирнов, 1976). Физиологический аспект проблемы широко разрабатывается в экспериментальных исследованиях различных лабораторий, результаты которых обобщены в монографиях и сборниках (Talland, 1965; John, 1967; Механизмы модуляции памяти, 1976; Механизмы управления памятью, 1979; Вартанян, Пирогов, 1987; Механизмы памяти, 1987, и др.). Стройная биоэлектрическая гипотеза памяти предложена М. Н. Ливановым (1977).
Данные о физиологических коррелятах памяти получены не только в эксперименте на животных, но и в исследованиях, проводимых у человека (Гречин, 1975; Раева, Ливанов, 1975; Кропотов, Гречин, 1976; John, 1977; Кропотов, 1979а, и др.).
Интерес к проблеме памяти все возрастает. При этом совершенствование методики исследований, формулирование и разработка новых концепций о механизмах мозга будут способствовать изучению процессов памяти. Широкие запросы педагогики и клиники остро ставят задачу изыскания путей управления процессом памяти, его различными свойствами. Поток обрушивающихся на человека сведений все увеличивается, поэтому возникла необходимость создать способы запоминания иногда очень большого количества сведений одновременно. И. М. Сеченов писал: «Память – сила, которая лежит в основе всего психического развития. Не будь в самом деле этой силы психическое развитие было бы невозможностью» (Сеченов, 1952а, с. 99).
Изыскивая предпосылки для улучшения запоминания, надо одновременно искать и противоядие – способы улучшения выделения значимости информации (эта задача в общем виде рассматривается техническими специалистами как проблема сигнал – шум), а также пути дезактивации, если не полного «стирания» памяти. Одновременная постановка этих задач важна и для обучения здорового мозга, и для переобучения мозга больного человека путем коррекции матрицы памяти, поддерживающей устойчивое патологическое состояние.
Обнаружение и исследование динамики физиологических изменений в различные фазы памяти оказываются возможными потому, что процесс запоминания в ходе обучения чаще всего происходит не мгновенно, а имеет развертку во времени. В то же время литературные данные и наблюдавшиеся нами при электрической стимуляции мозга явления типа импринтинга (запечатления) свидетельствуют о том, что мгновенное запоминание, запечатление возможно не только на самых ранних этапах онтогенеза (Бехтерева, Смирнов, 1975). Биологически, по-видимому, для организма выгодно иметь разные формы памяти. Реализация основных, важнейших для сохранения вида процессов организма и человека, и животных определяется генетической памятью, не развивающейся, а скорее проявляющейся в онтогенезе. Индивидуально приобретенная память определяет лишь адекватные коррекции протекающих процессов.
Для осуществления функций, важнейших для сохранения индивида как представителя вида, используется механизм мгновенной памяти – запечатление.
И, наконец, все другие виды деятельности, определяющие жизнь индивида в меняющейся среде и особенно жизнь человека в специально человеческой среде, базируются на более или менее быстро, но, как правило, не мгновенно формирующейся памяти. Скорость ее формирования является функцией значимости события и, таким образом, соответствующей мотивации. Она зависит от эмоционально активных или эмоционально нейтральных активационных механизмов и, следовательно, от функционального фона мозга. Этот процесс связан с генетическими особенностями мозга и целым рядом других, не всегда легко учитываемых факторов.
Биологической выгодой не мгновенного запоминания в процессе обучения является, по-видимому, возможность «отбора на запись», фильтрации информации и, что очень важно, одновременно идущего ее упорядочивания. В случае психонервной памяти примером упорядочивания является соотнесение ее с различными смысловыми и другими полями со всеми вытекающими отсюда последствиями иерархии дальнейшего ассоциативного поиска.
Генетически у человека имеются высокосовершенные предпосылки к психонервной памяти, реализующиеся в процессе обучения, индивидуального и особенно социально обогащенного развития. Так, не только обучение смысловому значению слов, но и использование их как важнейших единиц мыслительной деятельности осуществляется в ходе индивидуального развития человека в условиях общения с особями данного биологического вида. Именно в процессе обучения идет формирование и накопление базиса долговременной психонервной памяти при одновременной тренировке ее последующего использования в форме считывания, перевода из долгосрочной памяти в оперативную.
Как указывалось выше, связь мыслительных операций с процессами памяти неразрывна. Выделение в исследованиях собственно мыслительных процессов или собственно процесса памяти всегда имеет искусственный характер и определяет скорее расстановку акцентов, чем их сущность. Действительно, уже простейшие умозаключения немыслимы без активации и использования соответствующего базиса долгосрочной памяти вербального и невербального типов. В то же самое время по ходу тех же и всех других процессов все время происходит не только пополнение базиса памяти, но и трансформация ассоциаций и их иерархии. В полном соответствии со сказанным данные, полученные в процессе изучения тонких нейрофизиологических коррелятов слов и мыслительных процессов (Бехтерева и др., 1977а), оказались существенными для понимания целого ряда закономерностей нейропсихологии, нейролингвистики, физиологии анализаторных систем, искусственного интеллекта и ряда других направлений научных исследований. Они вместе с данными электрических воздействий на мозг оказались важнейшими для комплексно решаемой научной проблемы – проблемы памяти.
В предыдущей главе рассматривались результаты исследования перестроек импульсной активности, соотносимых с восприятием, удержанием в памяти и воспроизведением слов и некоторыми мыслительными операциями. Было показано, что в процессе восприятия слов в импульсной активности нейронных популяций формируется паттерн, характеризующийся изменениями частоты и структуры импульсного потока, формы разрядов нейронов и их групп внутри нейронных популяций и между дистантно расположенными популяциями. Уже в этом паттерне при использовании в психологических тестах слов, имеющих смысловую общность, обнаруживались нейрофизиологические корреляты этой общности в виде характерных перестроек взаимодействия нейронных группировок. Далее были выявлены перестройки структуры импульсного потока в виде очень динамичных определенных последовательностей интервалов и мало динамичных, но менее определенных изменений интервалов между импульсами. Общие перестройки структуры импульсных потоков, появляющиеся в разных зонах мозга (см. выше), рассматривались нами как фактор, способствующий организации мозговой системы. Общие последовательности интервалов, обнаруженные в коррелятах слов, соотносимых по смыслу, отражали переведенный из латентной формы в активную системообразующий фактор долгосрочной памяти. Они явились биоэлектрическим выражением активации ассоциативных связей, образовавшихся в результате индивидуального опыта и хранимых в долгосрочной памяти.
При инструкции запомнить слово и произнести его по команде и относительно небольших изменениях интервала между заданием и ответом можно видеть настраивание мозга на тот интервал в форме появления вначале слабо выраженных, а затем вполне отчетливых перестроек импульсной активности в виде появления паттерна сигнала, являющегося «оригинальной копией» первого развернутого паттерна, названного управляющим (Бехтерева и др., 1975б). Термин «оригинальная копия» заимствован из искусства. Если он и не очень удачен, то отражает и сходство управляющего паттерна с первым, возникшим при восприятии слова, и отличие от него. Отличие формально характеризуется его соотносимостью с акустическими (а вероятно, и моторными) характеристиками ответа больного. По существу, различие определяется тем, что второй паттерн в тестах с предъявлениями известных слов является результатом активации долгосрочной памяти лица, выполняющего тест. Подчеркивание в этих случаях значения активации долгосрочной памяти основано на сопоставлении нейрофизиологической динамики в тестах на краткосрочную память с динамикой при выполнении мыслительных операций. Если в первом случае соотношения относительно просты: полный первичный паттерн–компрессированный паттерн (паттерны)–полный управляющий паттерн, соотносимые с одним и тем же словом, то при различных входном и выходном сигналах, где входной является лишь одной из детерминант выходного – ответа, видно, как мозг формирует ответ при наличии соответствующего базиса долгосрочной памяти. Влияние последнего на динамику паттерна-кода в импульсной активности нейронных популяций прослеживается при варьировании наиболее простых тестов – тестов на краткосрочную память. При этом обнаруживается, что выполнение тестов, формально составленных одинаково, но с использованием слов, встречающихся в речи с разной частотой, известных и неизвестных, может осуществляться на различной нейрофизиологической основе.
Действительно, и при предъявлении редко и часто встречающихся в речи известных слов, и при предъявлении неизвестных слов по команде «Повторите» корректно или ошибочно воспроизводится то, что было в задании. Однако после восприятия известных слов наблюдается указанная выше нейрофизиологическая динамика, а при удержании в памяти неизвестного слова возникший при его восприятии паттерн сохраняется в импульсной активности, хотя и в дискретной форме, до речевого ответа. Сохранение паттерна связано в данных конкретных тестах и с тем, что больному предлагалось запомнить предъявленные сигналы. Однако идентичность этого условия при различии характера предъявляемых сигналов и соответственно этому различие мозговой нейродинамики позволили нам предположить, что второй вариант (сохранение паттерна) связан с отсутствием тормозящих влияний базиса долгосрочной памяти при неопознании субъективно нового сигнала. Нейрофизиологическим механизмом стабильности паттерна могут быть явления, протекающие по типу не до конца еще прослеженного и изученного процесса так называемой реверберации (Lorente de No, 1938; Беритов, 1960). Отношение циркуляции возбуждения по замкнутым нейронным цепочкам, спустя некоторое время после окончания действия какого-либо стимула к процессу краткосрочной памяти, недавно получило прямое экспериментальное доказательство на животных при исследовании взаимодействий функционально связанных нейронов (Вартанян и др., 1986). Стабильность, сохранение паттерна создают предпосылки к оптимизации в этих условиях формирования соответствующего биохимического базиса долгосрочной памяти. Это объяснение почти напрашивается, и мы пока не можем предложить лучшего, хотя, естественно, можно было бы предположить, что дело не в отсутствии тормозящих влияний, а в самом факторе новизны, активации соответствующих детекторов. Но что такое, если вдуматься в физиологическую сторону вопроса, новизна?
Зависимость обнаруженных во время удержания теста нейродинамических явлений от базиса долгосрочной памяти подтверждается не только различием их при разной степени субъективной новизны задания. Подтверждением является также и перестройка второго варианта нейродинамики на первый при обучении обследуемого лица значению ранее неизвестных иностранных слов. Тогда, когда звучание слова становится привычным, а значение – известным, тогда, когда можно думать, что соответствующий базис долгосрочной памяти сформировался, паттерн начинает исчезать из долгосрочной памяти так же, как это наблюдается при предъявлении известных слов. Данные, сопоставимые с этими нашими наблюдениями, уже встречаются в литературе. Об отражении в электрических явлениях мозга предыдущего опыта, считываемого из долгосрочной памяти, пишут Джон (John, 1976), В. А. Илюхина (1977) и другие авторы. Это неудивительно, так как не только в реальных жизненных условиях, но и в специально ориентированных физиологических экспериментах четко прослеживается влияние долгосрочной памяти на поведение, оценку внешних сигналов (Фирсов, 1977; Чораян, 1978, и многие другие).
Анализ составляющих паттернов-кодов и выделение динамичных групповых последовательностей разрядов с определенными интервалами позволили получить в тестах на обобщение новые, более детальные данные об отражении в импульсной активности нейронных популяций процесса активации долгосрочной памяти. После восприятия первых двух-трех слов одного смыслового поля в импульсной активности нейронных популяций появляется сложный фрагмент паттерна, в который входят и групповые последовательности разрядов, характерные для уже предъявленных слов, и новые элементы, позднее обнаруживаемые в паттерне-коде обобщающего слова (Гоголицын, 1976б). Этот феномен рассматривается как отражение уже происшедшего, хотя, возможно, и не окончательного («может быть»?) обобщения по минимуму слов смыслового поля. В плане рассматриваемой проблемы памяти он свидетельствует об извлечении обобщающего слова (естественно, в данном случае – активации его биоэлектрического паттерна) из долгосрочной памяти.
На примере процесса формирования паттерна-кода обобщения еще раз подчеркивается значение биоэлектрического компонента паттерна. Слово-обобщение («мебель», «деревья», «цветы» и т. д.) В заданиях отсутствовало. Оно проявляется как результат активации соответствующей матрицы долгосрочной памяти. При этом оказывается, что в биоэлектрическое отражение активации этой матрицы долгосрочной памяти включены элементарные составляющие биоэлектрических паттернов-кодов слов, образующих смысловое ноле. Паттерн-код обобщающего слова воспроизводит отдельные элементы паттерна-кода обобщаемых слов в сочетании со своим, специфическим, паттерном. Это может рассматриваться как нейрофизиологическое свидетельство правомочности представления, что обобщение не есть простая выборка значимых элементов паттернов обобщаемых слов и их суммация, а есть появление нового качества.
Предъявление зрительного изображения (первая детерминация ассоциативного поиска), сопровождаемое определенным словесным заданием (вторая детерминация – ограничение ассоциативного поиска), привело, как указывалось в наших предыдущих исследованиях (Бехтерева и др., 1977а), к появлению в импульсной активности целого ряда различных паттернов-кодов слов, имеющих видовую и индивидуальную смысловую связь с детерминированным изображением и заданным мыслительным процессом. Продолжение на новом уровне этих исследований, на время приостановленных в пользу получения статистически достоверных результатов, даст возможность, по-видимому, представить объективные свидетельства характера и результатов ассоциативного поиска в процессе интеллектуально-мнестической деятельности, а также выявить, как, на основе какого количества и рода информации, переведенной из потенциальной формы в активную, из долгосрочной памяти в оперативную, принимается то или иное решение.
Такие исследования с учетом приведенных в предыдущей главе ограничений, и прежде всего динамичности, неустойчивости паттернов и их элементов перспективны для понимания механизмов и нейрофизиологической основы сложнейших нарушений психических функций. Важно подчеркнуть, что уже первые результаты этих исследований оказались интересными для развития проблемы памяти. В этом случае, так же как и в тестах на краткосрочную память, обнаруживались и развернутые, и компрессированные паттерны. В развернутой форме появлялись паттерны-коды, связанные с вербализацией задания и ответа. Подавляющее большинство паттернов-кодов остальных слов выявлялось в компрессированной форме. Эти данные рассматривались нами выше в плане оценки условий осуществления мыслительных операций. Если взглянуть на них в аспекте проблемы памяти, придется допустить, что, по крайней мере, большой массив вербальной памяти хранится именно в экономичной, компрессированной, а возможно, и в связанной, комплексной форме.
Появление развернутых паттернов-кодов в форме управляющих паттернов-кодов при восприятии слов в тестах на краткосрочную память дает основание думать, что развернутый сигнал и механизм развертки теснейшим образом связаны с опознанием сигнала и вербализацией ответа. В более общей форме можно думать, что механизм развертки является важнейшим в процессе обучения, а механизм компрессии – в развитии мыслительной деятельности. Так же как и компрессия паттерна, развертка для формирования ответа не может осуществляться без участия информационно-специфической долгосрочной памяти. И в то же время не исключено, что реализации развертки могут принимать участие и другие, менее пецифичные механизмы мозга.
Не участвуют ли в процессах компрессии и развертки паттерна эмоционально активные и эмоционально нейтральные активирующие и тормозные влияния, значение которых и в мыслительных процессах, и в механизмах памяти хотя и общеизвестно, но далеко еще не понято? Развитие эмоционально активной или эмоционально нейтральной реакции активации или торможения в нейронных популяциях прежде всего влияет на текущую частоту разрядов. Исследования в этом направлении проводятся. Возможно, что они могут привести к переоценке физиологической природы компрессированных паттернов. До сих пор компрессированные паттерны по-прежнему (с 1971 года) рассматриваются как редуцированная форма развернутых, сохраняющая их опорные элементы. Не отрицая такой точки зрения, стоит рассмотреть вопрос, не являются ли компрессированные паттерны хотя бы частично результатом истинной компрессии элементов паттерна, чем-то вроде отражения компрессии шкалы времени. Как известно, вопрос о возможности жизни мозга по разному времени ставился в наших предыдущих исследованиях (Бехтерева, 1971, 1974, 1978). Он находит также подтверждения в психологических исследованиях (Брехман, 1976). Однако, как вполне понятно, для рассмотрения конкретных вышеприведенных данных в таком аспекте оснований пока еще недостаточно.
Результаты изучения кодирования мыслительных процессов дают все основания предполагать наличие в глубоких структурах мозга не только энергетических, но и информационноспецифических звеньев системы обеспечения психической деятельности. Однако сила традиционных представлений в науке о мозге человека обусловила первоначально объяснение нейрофизиологических явлений, наблюдавшихся местно в подкорке, как отражение событий на корковом уровне. Так, предполагалось, что где бы в мозгу ни возникал характерный биоэлектрический кодовый паттерн, адресом долгосрочной памяти была кора больших полушарий (Бехтерева, 1974). Надо сказать, аналогичная трактовка явлений, развивающихся на подкорковом уровне при психической деятельности, и сейчас еще очень распространена, хотя, как показано в предыдущей главе, накапливается все больше данных в пользу роли по крайней мере ряда подкорковых структур в процессах высшей интегративной деятельности.
Изучение вопроса заставило нас отказаться от этой предвзятости и вносить коррекции в гипотезу о пространственных соотношениях интеллектуальных процессов с их базисом – долгосрочной памятью.
Далее эти представления были нами развиты в форме гипотезы о распределенном кодировании вербальных сигналов. Нейрофизиологическим выражением этого распределенного кодирования было наличие преимущественно различных групповых и лишь отдельных высокосходных последовательностей разрядов в различных зонах мозга.
По-видимому, даже при исключении избыточности местных элементов паттерна-кода полный мозговой паттерн-код слова представляет собой интегральную величину, результирующую паттернов-кодов в различных звеньях системы обеспечения психической деятельности. Здесь важно подчеркнуть: слова «интегральная» и «результирующая» отражают сегодняшнее знание того, что полный код не есть простая сумма паттернов-кодов в различных нейронных популяциях и одновременно неполное понимание нейрофизиологических процессов, протекающих при объединении не трех, четырех, пяти, а многих звеньев систем. Выше уже рассматривались предположительные варианты структурно-функциональной организации мозга в этом случае.
Нейрофизиологические исследования показывают, что количество звеньев системы очень велико, особенно с учетом наличия в ней не только жестких, обязательных, но и гибких звеньев. Это положение подтверждается практически в каждом нейрофизиологическом исследовании мозга человека при использовании самых разных показателей жизнедеятельности мозга (Кропотов, Гречин, 1976; Кропотов, Малышев, 1976; Методы клинической нейрофизиологии, 1977; Кропотов, 1979а; Медведев, Белов, 1986, и др.). Так, воспроизводимые изменения при пробах на оперативную память наблюдались В. Б. Гречиным в 72 % точек мозга из общего числа 1000 обследованных зон в пределах премоторной коры, ядер таламуса и стриопаллидарной системы и верхних отделов ствола. При этом до 18 % обследованных зон обнаружили в большей или меньшей степени свойства детекции ошибки (зоны в хвостатом ядре, срединном центре, подушке, переднем и латеральных ядрах таламуса и др.). Выше показано, что в исследованиях, проведенных Ю. Д. Кропотовым и сотрудниками (Бехтерева и др., 1985б), данные о детекции ошибок получили полное подтверждение и дальнейшее развитие. Паттерн-код, индивидуальный в каждой нейронной популяции, огромное количество звеньев системы, казалось бы, создают свой неповторимый мозговой код у каждого обследуемого лица, что формально согласуется с данными психологии об индивидуальных психологических различиях разных лиц. И не только психологии. Электрическая стимуляция мозга человека обнаруживает также много индивидуального в его организации и особенно в том, что касается высших функций (Ojemann, 1979). Однако калейдоскоп групповых последовательностей разрядов в нейронных популяциях состоит из кодовых элементарных последовательностей, общие элементы которых обнаруживаются (по-видимому, играя роль системообразующего фактора) в разных нейронных популяциях в паттернах одного слова и в одной и той же популяции в паттернах-кодах слов со смысловой общностью. Следует здесь еще раз подчеркнуть уже упоминавшиеся выше данные Н. Г. Шкуриной (1983, 1984) о возможности наблюдения не только динамичных, но и сравнительно стабильных перестроек структуры импульсного потока, хотя и о значительно меньшей определенностью конкретных интервалов между импульсными разрядами нейронов.
Исследования механизмов и нейрофизиологических коррелятов общности явлений в разных звеньях мозговой системы, дальнейший анализ и систематизация кодовых паттернов будут, по-видимому, способствовать выяснению видовых характеристик (принципов!) отражения словесных сигналов в мозгу разных людей.
Приведение этих данных в настоящей главе было необходимо для анализа или, точнее, пересмотра представлений о месте и способе хранения информации в долгосрочной памяти, о локализации так называемых энграмм памяти. Распределенность кода при значимости его элементов, обнаруженных, в частности, в подкорковых нейронных популяциях, позволяет выдвинуть гипотезу о распределенном характере матрицы долгосрочной вербальной памяти и о значении подкорковых образований для распределенного хранения.
Исследуя форму нейронных разрядов при проведении тестов на краткосрочную память с предъявлением часто и редко встречающихся в речи известных слов и незначимых триграмм, П. В. Бундзен (Бехтерева и др., 1973) показал зависимую от степени известности слова и таким образом от базиса долгосрочной памяти динамику стабилизировавшихся по форме разрядов. Стабилизация по форме была кратковременной и появлялась при восприятии слова и его воспроизведении в случае предъявления часто встречающихся в речи известных слов, обнаруживалась более продолжительно в импульсной активности при предъявлении редко встречающихся известных слов и наблюдалась на протяжении всего удержания в памяти, причем с тенденцией распространения по пространству (популяции). На основе экспериментальных исследований (Rosenthal, 1967; Ходоров, 1969; Machek, Pavlik, 1973; Вислобоков, 1974) можно предположить, что это явление отражает определенное состояние мембран нервных клеток, предположительно соотносимое с процессами, лежащими в основе активации или формирования базиса долгосрочной памяти. Обнаружение этих явлений в подкорковых структурах подтверждает вероятность представлений о распределенном, не только корковом, но и подкорковом расположении (хранении) матриц долгосрочной памяти.
Если позволить себе некоторое отступление и вспомнить статью Лешли «В поисках энграмм» (Lashley, 1966), то, вероятно, именно распределенностью памяти хотя бы частично можно объяснить сложности, которые встречались и будут встречаться на пути поиска энграмм. Нет оснований представлять себе память структурно отделенной от деятельности, хотя опыт клиникофизиолого-анатомических сопоставлений и показывает особую значимость ряда образований мозга для нормального протекания процессов памяти. Если стоять на позициях распределенного хранения долгосрочной памяти, можно рассмотреть под определенным углом зрения затянувшиеся споры об уровнях замыкания условного рефлекса и об идентичности, и, наоборот, о различиях понятий «условный рефлекс» и «временна?
я связь». Формирование условного рефлекса основано на формировании соответствующего базиса долгосрочной памяти. С другой стороны, условный рефлекс есть форма выражения долгосрочной памяти. Долгосрочная память, не имеющая, по-видимому, строго определенного уровня формирования и хранения, а являющаяся функцией очень многих пространственно распределенных нервных клеток, и определяет нередко удивлявшую экспериментаторов возможность формирования пусть не классического условного рефлекса, а временно?
й связи при филогенетическом или хирургически детерминированном неучастии высших отделов мозга. С другой стороны, условный рефлекс, формирующийся у высших животных и у человека, протекающий при участии коры больших полушарий, нельзя себе представлять как процесс, где матрица памяти, ответственная за замыкание условной связи, имеет строго определенный, только корковый уровень. Даже самый простой условный рефлекс, затрагивая по ходу предъявления индифферентного (условного!) и безусловного сигналов разные уровни мозга, оставляет в них иногда, по-видимому, неизгладимые следы. Этих следов при длительном повторении одного и того же условного рефлекса может становиться меньше за счет процесса оптимальной минимизации системы, обеспечивающего деятельность и находящегося в известной мере в конкурентных отношениях с процессом распределения (не распределенности!) хранения памяти. Это приводит к тому, что даже для тех организмов, для которых задаваемая условно-рефлекторная деятельность является адекватной по сложности, любой условно-рефлекторный процесс при упрочивании начинает базироваться на распределенной, но оптимально минимизированной матрице памяти.
Переходящие из статьи в статью, из монографии в монографию данные Пенфильда о возможности наблюдать развернутые картины прошлого опыта при электрической стимуляции некоторых зон коры и глубоких отделов височных долей мозга, позволившие ему выдвинуть представления о коре и этих зонах, в частности, как о хранилище прошлого опыта и соответственно – о более глубоком структурном уровне высшей интегративной деятельности в мозгу, тоже, по-видимому, могут быть рассмотрены и с иных позиций. Если матрица долгосрочной памяти распределена, то при стимуляции коры и других зон эпилептического мозга, характеризующегося в определенные фазы болезни сочетанием исключительной проводимости и разных по локализации доминантных эпилептогенных очагов, могла быть активирована вся матрица, в которой хранилась память этого прошлого опыта, где бы ни находились ее звенья, по принципу, аналогичному ассоциативному поиску при мыслительных операциях.
То, что феномен воспроизведения прошлого опыта наблюдался при операциях по поводу эпилепсии при электрической стимуляции не только височной коры, но и медиобазальных отделов височной доли, нередко заставляло считать уже эти структуры хранилищами памяти. Имея в активе большой опыт стимуляции мозга разных больных, подтвердивший распределенность памяти (Вартанян, 1976; Смирнов, 1976; John, 1976; Ojemann, 1977a, 1978a; Смирнов, Бородкин, 1979), результаты электрической стимуляции коры и глубоких отделов мозга во время операций по поводу эпилепсии, по-видимому, можно рассматривать следующим образом. Они свидетельствуют о том, что стимулируемая зона коры или глубоких отделов мозга – значимый элемент матрицы памяти индивидуального опыта. Эти результаты, особенно если учесть свойства эпилептического мозга, не свидетельствуют в пользу положения, что кора или медиобазальные структуры височной доли являются единственными кладовыми, центрами этой памяти.
Распределенность памяти должна, несомненно, учитываться не только при изучении мозгового обеспечения мыслительных, но и всех других процессов здорового и больного мозга. Так, участие коры головного мозга в обеспечении основных процессов жизнедеятельности здорового и больного организма, показанное в трудах К. М. Быкова (Быков, 1947; Быков, Курцин, 1960) и его сотрудников, свидетельствует о существовании значимого в норме, искусственно сформированного у здорового организма или созданного болезнью коркового звена системы, имеющей, как известно, множество преимущественно подкорковых звеньев. Местный эпилептогенный очаг при эпилепсии, являющейся болезнью всего мозга и, по-видимому, организма в целом, есть закрепившаяся реакция мозга на поражение, местный элемент распределенной матрицы долгосрочной памяти. Такая трактовка позволила предположить в комплексном лечении целенаправленное воздействие, предположительно дезинтегрирующее именно память, обеспечивающую устойчивость этой патологической реакции. Результаты такого вмешательства достаточно обнадеживающие. Эпилептогенный очаг на ЭЭГ при формальной аналогии с процессами на нейронном уровне может быть сопоставлен с устойчивым паттерном-кодом на предъявление неизвестного слова, где устойчивость паттерна служила фактором, способствующим формированию базиса долгосрочной памяти в мозгу •больных паркинсонизмом. Более того, длительно существующий или воспроизводящийся в той же зоне мозга эпилептогенный очаг является уже и биоэлектрическим коррелятом долгосрочной памяти. При эпилепсии местный эпилептогенный очаг на ЭЭГ не только сформировал «память» данной реакции, но и трудности ее забывания, перевода в латентную форму, причем динамика биоэлектрической активности в зоне эпилептогенного очага свидетельствует о разных ее состояниях.
При анализе динамики паттерна-кода, наблюдавшейся при предъявлении хорошо известных слов, быстрое исчезновение паттерна-кода было отнесено за счет тормозящего влияния долгосрочной памяти. Как указывалось, торможение (угнетение) местного эпилептогенеза было достигнуто за счет воздействия, имитирующего биоэлектрические проявления важнейшего компонента сложного многопланового процесса памяти – забывания, дезинтеграции, угнетения (торможения) местной памяти о болезни.
Аналогия между данными нейрофизиологии психической деятельности и результатами нейрофизиологического изучения больного мозга была названа «формальной». И действительно, внешне это выглядит именно так. Однако длительное полиметодическое изучение здорового и больного мозга человека заставляет думать не только о чертах различия, но и об общности многих, внешне разных проявлений здорового и больного мозга и о трансформациях механизмов, исходно присущих здоровому мозгу, в патологические проявления. Очень важно, в том числе и в практических целях, уловить в патологических проявлениях отзвуки механизмов здорового мозга. Это станет возможным при более глубоком проникновении в законы деятельности и здорового, и больного мозга.
Распределенный характер хранения памяти обнаруживается в самых различных наблюдениях. Однако вопрос о том, какова роль различных образований мозга в процессах памяти, очень сложен. В подавляющем большинстве случаев вопрос должен и может анализироваться в связи с характером деятельности, общим функциональным состоянием мозга и его индивидуальным формированием. Именно в соответствии с этим и могут рассматриваться результаты лечебной экстирпации и точечной электрической стимуляции некоторых зон мозга человека.
Хотя в последние годы опубликованы данные, позволяющие предположить, что после разрушения медиобазальных отделов височных долей внешне проявляющиеся нарушения памяти, как, например, невозможность фиксации текущего опыта, связаны более с нарушением считывания, основные положения, показанные в эксперименте в конце 30-х годов (феномен Клювера– Бюси) и подтвержденные в клинике, остаются в силе. Разрушение этих областей мозга как бы прекращает возможность научения, хотя и не снижает заметно интеллектуальных функций, связанных с использованием прошлого опыта. При двухстороннем разрушении медиобазальных отделов височных долей больной разговаривает вполне осмысленно, производит впечатление интеллектуально сохранного человека, но стоит ему отвернуться или просто отвлечься чем-либо, как он вновь здоровается с тем, с кем разговаривал несколько минут назад. Он не может выполнить простейшего задания, если оно связано с удержанием в памяти последовательности даже нескольких операций. С течением времени утраченные способности восстанавливаются незначительно или не восстанавливаются совсем. Наличие сходной картины при склерозе сосудов мозга не противоречит представлению о ведущей роли медиобазальных структур мозга в этих расстройствах. Общее нарушение мозгового кровоснабжения может особенно сильно отразиться именно на этих структурах, ибо их кровоснабжение и в норме достаточно скудно, а заболевание лишь усугубляет проявления этой недостаточности. Важность медиобазальных отделов височных долей для функций памяти, а следовательно, и мыслительных функций требует уточнения стороны преимущественного поражения медиобазальных отделов височных долей у больных эпилепсией при решении вопроса об операции. Свойство эпилептического мозга формировать так называемые зеркальные очаги эпилептогенеза, имеющие несомненное отношение не только к процессам передачи информации в мозгу, но и к функции памяти, при длительно текущей эпилепсии определяет не просто желательность, а прямую необходимость использования долгосрочных вживленных электродов для уточнения стороны первичного или преимущественного поражения. Исходно билатеральное поражение медиобазальных отделов височных долей, в принципе, возможно, но встречается достаточно редко. Тщательное клиническое обследование с учетом нейрофизиологических и биохимических данных позволит решить, какая из двух симметричных зон способна к реабилитации в случае экстирпации одной из них с лечебными целями. Однако эти вопросы уже далеко выходят за рамки собственно проблемы памяти и, кроме того, специально рассмотрены в книге Н. П. Бехтеревой с соавторами (1978) и ряде других работ.
Полиметодический подход к исследованию механизмов мозга, преимущества которого были рассмотрены нами специально в первой главе, несомненно, должен быть использован и при изучении механизмов памяти. Наряду с данными анализа физиологических показателей жизнедеятельности мозга важным компонентом комплексного метода его изучения является, как известно, исследование результатов точечной электрической стимуляции.
К настоящему времени в результате стимуляции мозга при одномоментных стереотаксических операциях и в процессе диагностики и лечения методом вживленных электродов накоплен уже значительный материал не только о структурно-функциональной организации мозга, но и в отношепии изменения краткосрочной памяти и процессов считывания из долгосрочной в зависимости от области и характера воздействия, более или менее жестко связанных с изменениями состояния и других функций мозга (Смирнов, 1976; Ojemann, 1977b, 1978a, и др.).
При электрической стимуляции гиппокампа или его извилины у больных эпилепсией наблюдались явления типа антеро– и ретроградной амнезии. Они продолжались от нескольких секунд до нескольких недель в зависимости от интенсивности и длительности стимуляции. Больной в эти секунды или недели может забыть то, что непосредственно предшествовало воздействию, и не запоминает текущих событий, долговременная же память не страдает, а непосредственное воспроизведение заданий иногда даже улучшается. Полиметодический подход к оценке физиологических коррелятов развивающихся нарушений памяти и появление их одновременно с признаками функционального выключения мозга (разряд после действия на ЭЭГ) объясняют сходством явлений, имевших место при экстирпации и некоторых видах стимуляции мозга, свидетельствуют о целесообразности отнесения наблюдавшихся дефектов памяти именно за счет выключения данных медиобазальных образований височной доли и лишний раз подчеркивают необходимость осторожности при физиологической оценке эффектов так называемого стимулирующего воздействия.
Преходящие, слабо выраженные изменения памяти наблюдались при стимуляции передних отделов поясной извилины (Whitty, Lewin, 1960; Fedio, Ommaya, 1970). Восприятие (запоминание) невербальных сигналов было затруднено при стимуляции правого мамиллоталамического тракта (Ojemann, 1971). Отсроченное непостоянное улучшение вербальной памяти имело место после лечебных электрических стимуляций коры мозжечка (Riklan et al., 1976). Эти изменения наблюдались одновременно с нормализацией эмоционально-мотивационного состояния больных и рассматривались как вторичные по отношению к данным эмоциональным сдвигам и подавлению активности эпилептогенного очага.
Специальное исследование памяти и внимания у больных эпилепсией предприняли в нашей лаборатории А. Н. Шандурина и Г. В. Калягина (1979) при электростимуляции гиппокампа, его извилины, септальных ядер свода, передней комиссуры, срединного центра, интраламинарных, вентролатеральных и передневентральных ядер таламуса, мезэнцефальной ретикулярной формации и некоторых зон неокортекса. Исследование показало, что под влиянием электростимуляции, независимо от точки приложения тока, наибольшей изменчивостью обладала вербальная и невербальная долговременная память, устойчивость и переключаемость внимания. При электростимуляции структур правого полушария несколько больше изменялась динамика объема кратковременной образной памяти, левого – объема кратковременной памяти на цифры. При этом правосторонние воздействия чаще вызывали динамику объема внимания, левосторонние – его устойчивости. Электростимуляция таламических ядер и мезэнцефальной ретикулярной формации вызывала обычно одновременное изменение кратковременной и долговременной вербальной и образной памяти, сочетавшееся с изменениями внимания. Изменения отдельных видов памяти, часто (но не всегда) не связанные с динамикой объема внимания, наблюдались при электрической стимуляции медиобазальных образований левой височной доли. При этом могли обнаруживаться одновременные изменения кратковременной и долговременной образной памяти, кратковременной зрительной вербальной памяти, в последнем случае при одновременном изменении объема внимания. Изменения и внимания, и памяти были различными при стимуляции медиобазальных структур височной доли и срединных структур мозга. При электростимуляции таламических и мезэнцефальных структур чаще изменялся объем внимания и слуховая кратковременная память, а при электростимуляции медиобазальных отделов височных долей – устойчивость и переключаемость внимания и вербальная и образная зрительная кратковременная память. В исследовании показано дифференцированное влияние электростимуляции различных образований медиобазальных отделов височной доли, в частности, гиппокампа и миндалины, на динамику мнестических функций разных модальностей.
Специально исследовалась динамика фаз мнестической функции при электростимуляции. Электростимуляция ядер таламуса чаще вызывала изменения объема кратковременной вербальной памяти, а при электростимуляции медиобазальных образований – изменения отсроченного воспроизведения заученного до воздействия вербального материала. Следует подчеркнуть, что детальное изучение изменений памяти при электростимуляциях вышеуказанных структур выявило и другие изменения памяти, в частности, в форме ее дезинтеграции. 30 % электрических стимуляций этих образований сопровождались изменениями речи в виде ускорения или замедления ее темпа, дизартрии, аномии и др., 20 % – изменениями «схемы тела».
Не детализируя специально этого вопроса, авторы указывают, что электростимуляция редкими импульсами (3–6 Гц), собственно стимулирующее воздействие на миндалину вызывали улучшение зрительной вербальной памяти при повышении показателей психической деятельности.
Изучение эффектов не только отдельной электростимуляции, но и результатов курса лечебных стимуляций неспецифических таламических ядер показало, что улучшение кратковременной и долговременной памяти отмечается наряду с общей нормализацией психических функций. Положительная динамика этих функций, в свою очередь, коррелировала с уменьшением проявлений эпилептогенеза. В работах Мильнер (Milner, 1970), А. Р. Лурия (1974), Н. К. Киященко с соавторами (1975) широко представлена гипотеза о модально неспецифических влияниях подкорковых структур мозга на память.
Данные А. Н. Шандуриной и Г. В. Калягиной (1979) свидетельствуют о модально-специфическом влиянии одних и модально-неспецифическом – других образований мозга на краткосрочную память. Так, слуховые виды краткосрочной памяти чаще изменяются при электростимуляции таламических и мезэнцефальных структур, зрительных, медиобазальных отделов височной доли. Изменения памяти при стимуляции гиппокампа были модальнонеспецифическими, и в то же время именно эта структура во многом определяла состояние функции памяти. Ключ к пониманию причин различия указанных выше результатов – в методике исследования. Как известно, многие исследователи в области нейропсихологии основываются в своих построениях на динамике психических функций при массивных поражениях мозга. Точечные электрические стимуляции, одномоментные или в виде целого курса лечебных стимуляций, позволяют получить значительно более тонкие данные о структурно-функциональной организации мозга и, в том числе о мозговой организации памяти. Изменения памяти при точечной электрической стимуляции глубоких структур мозга рассмотрены в ряде монографических работ. Значительные изменения краткосрочной памяти в этих условиях наблюдал В. М. Смирнов (1976). Им описаны сверхоптимальные состояния, при которых наряду с общей психической активацией может почти вдвое (!) увеличиваться объем краткосрочной памяти. Такого рода эффекты отмечены, в частности, при электрической стимуляции срединного центра зрительного бугра. В. М. Смирнов указывает также, что по количеству зон, точечная электрическая стимуляция которых вызывала изменения памяти, на первом месте стоят ядра таламуса – подушечное и заднелатеральное. Значительное количество точек, электростимуляция которых вела к изменению краткосрочной памяти, обнаружено в области вентролатерального ядра и срединного центра зрительного бугра и бледного шара, а также мезэнцефального отдела ствола. В. М. Смирнов подчеркивает, что эффекты электрической стимуляции этих зон мозга на краткосрочную память существенно зависели от общего функционального фона. В соответствии с этим они могли варьировать в случае повторной стимуляции той же зоны даже при использовании тех же характеристик тока или не проявляться совсем.
Для оценки физиологической сущности эффекта, развивающегося при электрической стимуляции различных зон мозга, важно было учитывать характеристики используемых электрических воздействий. Эта задача облегчалась тем, что в процессе диагностики и лечения методом вживленных электродов не только использовалась стимуляция короткими (1 мс) прямоугольными импульсами различной частоты, но и применялась точечная поляризация постоянным током до 1 мА, приводящая к временному местному угнетению функций мозга. Сравнительный анализ данных показал, что эффекты электрической стимуляции частотой 50 имп./с приближались к явлениям, характерным для микрополяризации. Следовательно, истинный эффект стимуляции по крайней мере ряда мозговых зон наблюдается только при воздействии редкими импульсами – 4, 8, 10 имп./с. Увеличение объема краткосрочной памяти при электрической стимуляции бледного шара и некоторых зон мезэнцефальных отделов ствола с частотой 50 имп./с дает, следовательно, основания полагать, что в пределах указанных образований мозга имеются зоны, в естественных условиях тормозящие эти процессы.
Многолетние исследования памяти при электрической стимуляции мозга отражены в трудах Оджеманна (Ojemann, Fedio, 1968; Ojemann, 1978а). В работах 1968 года он показывает, что электрическая стимуляция подушки зрительного бугра может привести к невозможности или ошибочности называния предмета при сохранности экспрессивной речи и забывании изображения, предъявленного непосредственно перед стимуляцией. Используя методику изучения вербальной памяти с одновременным отвлечением внимания, Оджеманн (Ojemann, 1978a) исследовал влияние электрической стимуляции вентролатерального ядра таламуса (у больных паркинсонизмом во время стереотаксических операций) и коры (во время краниотомии с целью иссечения эпилептогенных очагов у больных эпилепсией).
Электрическая стимуляция проводилась на фоне предъявления стимулов, помехи (отвлечения внимания) или в момент активации словесной командой долгосрочной памяти: предлагалось назвать предъявленный на слайде предмет. Статистически значимые различия краткосрочной вербальной памяти обнаружены при стимуляции левосторонних ядер зрительного бугра. Стимуляция вызывала уменьшение числа ошибок в ответе, если совпадала с заданием, и увеличение – если совпадала с ответом. Стимуляция во время отвлечения внимания на правильность ответа не влияла. Одновременно было отмечено, что стимуляция левого таламуса вызывает специфическую реакцию настораживания, которая фиксирует внимание на предъявляемой вербальной информации. Исследования долгосрочной памяти у больных после операции показали, что стимуляция левосторонних ядер зрительного бугра влияет не только на краткосрочную память, но и на долгосрочную.
Большой интерес представляет описанное Оджеманном уменьшение аномии, возникшей при стимуляции левого вентролатерального ядра у больного после инсульта за несколько лет до стереотаксической операции, а также предположение об участии левого вентролатерального таламуса в обеспечении смысловой памяти. При стимуляции медиальной центральной зоны левого вентролатерального ядра таламуса правильное называние предметов нарушалось, причем зона ядра, стимуляция которой приводила к нарушению данного вида долгосрочной памяти, была территориально меньше зоны, стимуляция которой влияла на краткосрочную память. Наряду с этими данными было показано влияние левого вентролатерального ядра на речевую функцию (Ojemann, Ward, 1971; Ojemann, 1977a). При этом области, стимуляция которых влияла на память и речь, перекрывали друг друга, имели и общие и различные зоны (заднелобная, теменная и височная кора у больных эпилепсией). Нарушения называния предметов возникли при стимуляции левой теменно-височной области. Изменения краткосрочной памяти выявлялись при стимуляции зоны теменной и височной коры, прилежащей к речевой области.
О возможности наблюдать изменения памяти при электрической стимуляции тех же зон мозга, которые использовались в качестве лечебных мишеней, пишет и Сем-Якобсен (Sem-Jacobsen, 1968). Он указывает, что в этих условиях возможны явления типа воспроизведения прошлого опыта (подобно тем, которые наблюдал Пенфильд) при стимуляции правой височной доли и области слева от третьего желудочка, передних и задних отделов лобных долей. Очень детально анализируя все эффекты стимуляции, Сем-Якобсен подчеркивает возможность одновременного наблюдения как изолированных реакций, так и их комплекса, в котором изменения памяти сочетались с изменениями речи, настроения, сознания, сердечно-сосудистыми и другими вегетативными реакциями. Эффекты в наблюдениях Сем-Якобсена были воспроизводимыми при повторных стимуляциях тех же зон.
Обобщая опыт исследования различных реакций мозга человека и животных во время точечных электрических стимуляций, Валенштейн (Valenstain, 1973) приходит к важному заключению в отношении, прежде всего, вызванных эмоционально-психических реакций. Видовое сходство анатомического строения мозга не является предпосылкой сходства реакций у разных больных при стимуляции одной и той же анатомической зоны. Что же касается эффектов стимуляции, которые могут расцениваться как связанные с нарушением, активацией или подавлением памяти, то они, по впечатлениям Валенштейна, не воспроизводятся жестко даже у одного и того же больного, а не только у разных больных. При этом Валенштейн рассматривает критически утверждения, по крайней мере, одной группы авторов – Пенфильда и его сотрудников, настаивавших на полной воспроизводимости картин прошлого опыта. Кстати, несмотря на то что по существу Валенштейн, повидимому, прав, его сомнения относительно этих эффектов могут быть проверены лишь в дальнейших исследованиях. Валенштейн полагает, что авторы, настаивавшие на их воспроизводимости, были под гипнозом представлений Зигмунда Фрейда, бо?льшая роль в которых отводится «подавленной памяти о прошлом».
Целенаправленный анализ данных, приводимых Валенштейном, Сем-Якобсеном, Пенфильдом и другими исследователями, показывает, что многие разночтения данных связаны с особенностями методических решений. Так, в работах Пенфильда полная воспроизводимость картин прошлого опыта при стимуляции одной и той же зоны мозга отмечалась при частых стимуляциях. После перерыва в стимуляциях эффект мог возникнуть, но быть отличным от предыдущих. Картина, извлекаемая из прошлого опыта, могла оказаться другой, чем после первых стимуляций. Такие же изменения эффектов во времени, но уже при стимуляции через вживленные электроды наблюдали Стивенс с соавторами (Stevens et al., 1969). Кстати, эту динамичность эффектов интересно и важно, по-видимому, в дальнейшем сопоставить с динамичностью биоэлектрических паттернов-кодов.
Изучение влияний на процессы памяти различных мозговых зон важно само по себе как раздел фундаментальных исследований мозга человека. Оно имеет несомненное значение не только для понимания нормальной психической деятельности и различных проявлений нарушений психонервной памяти в клинике, но и для понимания основы устойчивости патологических состояний при болезнях мозга. И уже сейчас эта проблема важна потому, что, изучая, как именно память влияет на самые различные процессы, можно и нужно разрабатывать на этой основе пути и приемы воздействий, включающих как основной или дополнительный элемент направленную модуляцию процессов памяти.
Что же иллюстрируют приведенные хотя и не полные, но полученные с помощью разных подходов, а отсюда репрезентативные данные о мозговой организации памяти человека? Прежде всего, несомненно, что, хотя существуют зоны мозга, имеющие очень тесную связь с процессами памяти, данные записи физиологических показателей мозга и его электрической стимуляции свидетельствуют об организации памяти по распределенному принципу. Самые разные структуры и зоны этих структур имеют отношение к памяти, причем то обстоятельство, что связь ряда образований мозга с памятью не выявлена, может зависеть и от методической стороны исследования, и от направленности интересов исследователей. Создается впечатление не просто о системном характере организации памяти, а о множестве систем, обеспечивающих различные виды и различные фазы памяти, имеющих общие для всех и различные для каждой из них звенья. Так, несомненно важное значение для памяти имеют медиобазальные отделы височной доли, причем, если отвлечься от того, что больной с двусторонним повреждением этих областей мозга при общении кажется интеллектуально сохранным, можно говорить об этих зонах мозга как важнейших для памяти. Но что значит – кажется интеллектуально сохранным? Это значит, прежде всего, что он владеет индивидуально сформированным и хранящимся в мозгу базисом памяти. Страдает при поражении медиобазальных отделов мозга перевод информации на долгосрочное хранение. Что же, долгосрочная память совсем не подвластна воздействиям? Весь опыт клиники диффузных и прежде всего сосудистых поражений мозга показывает, что и долгосрочная память может существенно нарушаться, или, точнее, нарушается возможность пользования всем этим богатством – считывания из долгосрочной памяти.
Наряду с этим, часто трудно дифференцируемым симптомокомплексом нарушений памяти, данные нейрофизиологических исследований представляют косвенные свидетельства местной активации долгосрочной памяти, почти позволяют «дотронуться» до нее, а воздействие путем электрической стимуляции на некоторые зоны мозга может ее избирательно нарушать. Пересмотр представлений о принципах хранения памяти в мозгу, введение концепции о распределенном ее хранении позволяют непредвзято оценить и эти данные. Стимуляция небольшой зоны в области зрительного бугра вызывает невозможность или ошибочность называния предметов. Долгосрочная память и обязательно механизм считывания из нее страдают. Следовательно, именно эти зоны можно отнести к образованиям мозга, являющимся очень важными для процессов долгосрочной памяти и по крайней мере пользования ею. Стимуляция конвекситальной коры приводила к считыванию из долгосрочной памяти картин прошлого опыта. Но, впрочем, именно эти факты нами уже обсуждались в самых разных аспектах. И далее, наряду с зонами мозга, влияющими на процессы памяти, независимо от ее модальности, описываются зоны, оказывающие на память и модально-специфическое влияние.
Стереотаксическая неврология еще только формируется. Ее дальнейшее развитие приведет, очевидно, по крайней мере к тому, что будут описаны достаточно полно не отдельные реакции, а спектры (синдромы!) реакций, развивающихся при точечных электрических воздействиях. Однако и сейчас не вызывает сомнений положение о том, что зоны, нейронно-глиальные популяции мозга, имеющие отношение к памяти, как правило, связаны – какими-то видами функций, хотя, по-видимому, есть зоны, при стимуляции которых влияние на память является доминирующим. Рассматривая все то, что приведено в настоящей книге, и то, что оставлено за ее пределами для сохранения логики изложения, для того чтобы за деревьями не потерять леса, невольно напрашивается мысль, что память не только очень общее свойство мозга, но и свойство всего мозга. Но для этого, демобилизующего дальнейший поиск вывода сейчас еще, к счастью, нет достаточных оснований (см. данные Ю. Д. Кропотова), а, судя по результатам изучения других базисных механизмов мозга, дело почти наверняка обстоит и не так сложно, и не так просто (рис. 36).
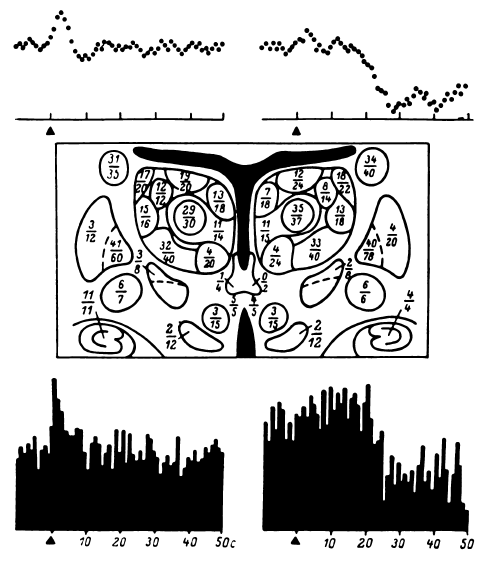
Рис. 36. Результаты статистической обработки тестов по запоминанию цифр у 40 больных паркинсонизмом.
Наверху – два типа динамики импеданса в двух различных структурах мозга (приведены характерные кривые). Внизу – два типа динамики мультиклеточной активности (постстимульные гистограммы). 1-й тип – изменения возникают в начале теста, 2-й тип – отсроченные изменения активности. В центре – на срезе мозга указаны структуры, в которых производились наблюдения. Цифры в знаменателе – общее число исследованных в данной структуре точек мозга, в числителе – число точек мозга, в которых наблюдались воспроизводимые изменения наблюдаемых физиологических процессов.
Что очень важно подчеркнуть сейчас? Кроме связи памяти со структурами мозга очень важным является усиление или ослабление этой связи в зависимости от местного и общего функционального состояния здорового и больного мозга. Процесс памяти, если и организованный в мозгу по полисистемному принципу, характеризуется наличием не только общих, ключевых звеньев для всех систем, но и звеньев более или менее жестких в каждой из систем. И хотя здесь, по-видимому, придется говорить о разных степенях жесткости, не исключено, что в основе положительных клинических эффектов разрушения, в частности, вентролатерального ядра при гиперкинезах (паркинсонизме) лежит не только выключение гиперактивного звена патологической системы и не только перерыв путей, но и выключение важного звена, влияющего на долгосрочную память! В то же время, если по крайней мере ряд звеньев системы (систем!) памяти жестко генетически структурно запрограммирован у человека как представителя вида, в мозгу каждого человека территориальная организация зон, модулирующих память, индивидуальна, а также преимущественно связана (точнее сцеплена) с деятельностью. Так, вентролатеральное ядро зрительного бугра, стимуляция которого влияет и на краткосрочную, и на долгосрочную память, является важным, хотя и компенсируемым при разрушении (гибким?) звеном системы обеспечения движений, эмоций, некоторых видов психической деятельности и т. д. А данные этого рода, в свою очередь, подтверждают высказанное в предыдущей главе положение о местных функциональных перестройках структур, связанных с различными видами деятельности и одновременно с записью, хранением и считыванием памяти.
Было бы неправильным ставить память наравне с другими функциями мозга. Память является базисным механизмом, свойством мозга, и факт ее организации также, по-видимому, по системному принципу не должен как бы низводить ее на уровень других систем. Память – тот основополагающий механизм, который лежит в основе возможности проявления всех онтогенетически формирующихся видов деятельности и соответственно всех обеспечивающих их мозговых систем. И в то же время во всех или во многих звеньях этих мозговых систем присутствуют базисные фиксирующие механизмы, позволяющие формироваться не только системе обеспечения функций, но одновременно и поддерживающей ее матрице памяти. Индивидуальность зон мозга, влияющих на память, существует, но может быть несколько преувеличена. Она может быть связана с индивидуальностью в строении мозга, которая, как известно, приводит к тому, что самые точные стереотаксические расчеты не в состоянии обеспечить применительно к расположению конкретных нейронноглиальных популяций полную сравнимость расположения в мозгу электрода при одномоментных стереотаксических операциях и при использовании метода вживленных электродов. В соответствии с этим уже даже факт получения однотипных результатов при стимуляции предположительно одной и той же зоны мозга, скажем, у двух-трех из десяти больных может расцениваться в пользу наличия не только индивидуальных, но и видовых закономерностей. Однако и исследования по изучению эффектов электрической стимуляции, и исследования нейрофизиологического кодирования психических процессов в мозгу свидетельствуют о значимости индивидуального паттерна (здесь – в широком смысле слова), накладывающегося на видовую принципиальную структурно-функциональную организацию мозга. Если попытаться проанализировать соотношение видового (надвидового) и индивидуального в механизмах мозга, можно было бы построить схему-шкалу: от видового к индивидуально-видовому, от индивидуально-видового к видовому – надвидовому. На этой схеме-шкале к видовому должно быть отнесено само строение мозга, его жесткая структурно-функционально генетически детерминированная организация. Индивидуально-видовой является организация индивидуально формирующихся систем деятельности и нейрофизиологических кодов психической деятельности, а вновь видовыми и надвидовыми – многие, если не все, биохимические и молекулярно-биологические основы функционирования мозга. Значение среднего звена схемы очень велико. Оно лежит в основе индивидуальности, неповторимости индивидуальных возможностей каждого человека. Значение правого звена схемы не только в общем положении единства человека со всеми живыми организмами. Оно и в других, более частных, положениях. Так, например, если данные о строении человеческого мозга могут быть получены только при изучении человека, а эволюционный подход способствует пониманию происхождения особенностей, присущих мозгу этого высшего млекопитающего, если данные о мозговой организации обеспечения индивидуально формирующихся у человека функций и, особенно специально человеческих, возможно получить только при исследовании, проводимом у человека, – при сугубо вспомогательном значении эксперимента, то изучение биохимических и молекулярно-биологических механизмов мозга вполне осуществимо в эксперименте на животных, в том числе и на сравнительно низко организованных. Правда, не исключено, что при исследовании мозга человека будут вскрыты новые стороны вопроса. Именно в данной работе обязательного упоминания заслуживают надвидовые свойства специфических полипептидов, свойства, позволяющие переносить патологические процессы (мозговую память о них?) от человека к животным (Вартанян, Балабанов, 1978) и, может быть, в дальнейшем – корригировать патологическую матрицу памяти человека полипептидами животного происхождения. Приведенные данные о мозговой организации памяти, ее структурном соотношении с деятельностью здорового мозга, лежащем в основе формирования матриц памяти, имеют значение для понимания законов функционирования больного мозга, многоплановых соотношений памяти и болезни.
Хроническое заболевание мозга формирует свой, новый рисунок матрицы памяти, перестраивая не только матрицу в целом, но и меняя удельный вес разных ее звеньев, создавая на самой основе закрепления реакций в памяти важнейшие звенья матрицы патологической памяти. Вероятно, излишне напоминать, что речь здесь идет не только и не столько о психонервной памяти, сколько о памяти как о базисном механизме, явлении и процессе мозга. Болезнь, таким образом, влияет на процессы памяти. В больном мозгу в связи с болезненными утратами и избыточной активностью формируется по тем же принципам, что и в норме, распределенная матрица, поддерживающая устойчивое патологическое состояние, влияющая далее на болезнь также по тем же основным законам, как и память здорового организма влияет на его функции. Так, например, стойкий эпилептогенный очаг, сформировавшись под влиянием постоянного раздражения в эпилептическом мозгу, становится далее важнейшим звеном матрицы памяти, поддерживающей устойчивое патологическое состояние, определяющим в большей мере и общее функциональное состояние мозга, и психонервную память, и эмоционально-психическую деятельность. В лечении хронических болезней мозга приходится использовать приемы воздействия на исходную вредность, на болезненную избыточность и недостаточность активности разных систем и структур, на матрицу памяти и отдельные ее звенья. Способ активного преодоления устойчивого патологического состояния и поддерживающей его матрицы долгосрочной памяти с помощью лечебных точечных электрических стимуляций мозга, предложенный впервые в нашей лаборатории В. М. Смирновым (Смирнов, Сперанский, 1972), затем примененный в Мадриде Дельгадо (Delgado, 1973), был возведен в ранг программных проблем конгрессов по стереотаксической и функциональной нейрохирургии (1979, июль, Париж). Прием для «стирания» памяти о местном эпилептогенезе рассмотрен выше. Успешное лечение фантомно-болевого синдрома оказывается возможным воздействием не только на проводящие болевые импульсы пути, но и на память о болезни: речь идет о наблюдавшейся нами дезинтеграции фантомно-болевого синдрома при лечебной электрической стимуляции подушки таламуса – ядра, активного в отношении памяти и не имеющего прямого отношения к самой боли.
Таким образом, исследование механизмов памяти здорового и больного мозга, вскрыв некоторые общие закономерности, открыло новые перспективы модуляции памяти в интересах больного и больных, еще далеко не до конца используемые. В то же самое время результаты исследования памяти здорового и больного мозга выявили общность принципиальных механизмов здорового и больного мозга, показали возможность понимания механизмов болезни на основе изучения механизмов здорового и целенаправленного управления механизмами больного мозга.



![Всё, что нужно знать о сахарном диабете [Незаменимая книга для диабетика]](https://img-lib.med-tutorial.ru/2970328225/cover.jpg)


