Некоторые положения в развитии проблемы. Факты и концепции
С позиций сегодняшних методических возможностей искать ошибки прошлого легко. Но практичнее для взгляда в будущее вылавливать в прошлом то, что прошло апробацию временем, – своего рода жемчужины, и отслеживать динамику формирования сокровищницы знаний о мозге. Laycock (цит. по: Jackson, 1931), И. М. Сеченов, В. М. Бехтерев, Дж. Джексон и многие другие ученые XIX и начала XX века не имели сегодняшних методов познания мозга. И все же, по крайней мере, два основных намеченных ими пути в проблеме «Мозг и психика» значимы и сейчас. Ими оказываются функциональная анатомия мозга человека и рефлекторный, а затем условно-рефлекторный принципы. У классиков XIX века были, как известно, и свои предшественники. Уже Аристотель придавал особое значение передним отделам мозга – в частности, «переднему» желудочку – значение высшей структуры мозга. В задачи этой работы не входит упоминание справедливо достойных имен науки по дороге длиной почти в два тысячелетия – от Аристотеля до взлета науки в XIX веке. Известно, однако, что в глубине прошедших веков было много мыслителей в близкой нам области науки, наиболее ярко представленных в книгах М. А. В. Brazier (например, 1984, и др.).
Основные вехи в развитии функциональной анатомии мыслительных процессов
Функциональная анатомия ведет свое начало с работ П. Брока (Вгоса, 1861). Строилась она первоначально на основе клинико-анатомических параллелей. Ее история хорошо известна, приведена во многих публикациях, и в том числе моих (Бехтерева, 1971, и др.). Существенный вклад в изучение функциональной организации мозга внесли наблюдения результатов электростимуляции мозга (Penfield, Jasper, 1954; Valenstein, 1973; Смирнов, 1976; Ojemann, 1983, и др.). В этих исследованиях была показана разница в функциональной организации коры и подкорковых структур. Электростимуляция коры височной доли могла вызывать появление связных картин прошлого опыта, ощущение раздвоения личности. Стимуляция подкорки приводила к развитию более элементарных реакций. Использование нативной электроэнцефалограммы человека очень мало дало для исследования мозговой организации высшей нервной деятельности (Gastaut et al., 1957). С развитием техники анализа вновь делаются попытки использовать электроэнцефалограмму для рассмотрения физиологических процессов, развивающихся в мозгу человека при мыслительной и эмоциональной деятельности (Ильюченок, 1996; Свидерская, Королькова, 1996; Яковенко, Черемушкин, 1996, и др.). ЭЭГ начинает занимать место все более достойного партнера в brainmapping, и прежде всего в связи с ее динамичностью. Происходит своего рода «возвращение» к электроэнцефалографии, к ее функциональным возможностям (Orrison et al., 1995; см. также: Neuroscan News. 1997. Vol. VI, N2).
Огромна литература по вызванным потенциалам (ВП). С помощью этой ручной и надежной методики получено множество ценнейших данных об обработке мозгом физических и смысловых характеристик речевых сигналов и, что особенно важно, об общих и частных механизмах мозга. А. М. Иваницким в исследованиях ВП (Ivanitsky, Strelets, 1977; Иваницкий, Стрелец, Корсаков, 1984; Иваницкий, 1996) была высказана идея, что субъективные феномены возникают в результате повторного входа возбуждения в места первичных проекций и сопоставления на этой основе новой информации с прошлым опытом. Сравнивая в одном и том же эксперименте амплитуду волн ВП с интенсивностью и временем ощущений, он предположил, что ощущение возникает в результате возврата возбуждения в проекционную кору после прохождения его через ассоциативную кору, гиппокамп и мотивационные подкорковые центры. Благодаря этому на нейронах проекционной коры происходит синтез сенсорной информации и сведений, извлекаемых из памяти. Момент этого синтеза с высокой точностью совпал с возникновением ощущений (как зрительных, так и соматосенсорных).
Позднее концепция возврата возбуждения в нервные структуры (англ. reentering) как основа осознаваемых феноменов была высказана рядом других авторов (Edelman, 1989; Stroering, Branch, 1993, и др.).
В последующие годы идею информационного синтеза как основы психических функций развил А. М. Иваницкий (Ivanitsky, 1993; Иваницкий, 1996) применительно к механизмам мышления. Сейчас с помощью ВП определяется ведущая зона во множестве активированных на ПЭТ-изображениях участков мозга и строятся различные представления о регулирующих и исполнительных системах мозга (Кропотов, Пономарев, 1993). Вызванные потенциалы – один из важных инструментов изучения мозговой организации самых различных процессов и прежде всего психических.
Сравнительно недавно (в календарном аспекте) пришло представление о возможности большой свободы индивидуальной функциональной организации мозга. Эта свобода максимально проявляется именно в организации мышления. Жесткими, с запрограммированными пространственными положениями звеньев в мозгу изначально, до рождения, являются системы обеспечения лишь сравнительно немногих функций – да и то, по крайней мере у некоторых из них, запрограммированность до рождения касается лишь расположения основной мозговой зоны. По ходу индивидуального развития идет торможение активности распространенных по мозгу функционально близких участков; такая схема приложима, в частности, для индивидуальной эволюции мозгового контроля движений.
Сходный, но не идентичный процесс происходит при формировании и закреплении мозговой системы обеспечения речи. Неидентичный – так как многие зоны мозга, лежащие вне классических речевых, остаются реально или потенциально активными. Для восстановления утраченной в результате травмы или сосудистого процесса функции речи их искусственная активация (электро– или магнитостимуляция) может оказаться очень и очень полезной. В моих работах (Бехтерева, 1988а; 1994а, б и др.) подробно описано, как это происходит в клинике.
Что касается менее автоматизированных мозговых функций, то там индивидуальное развитие лишь в некотором типовом варианте формирует звенья мозговых систем, обеспечивает множество возможных систем для выполнения каждой функции, причем в большой зависимости от того, в каких условиях эта деятельность осуществляется. На срезах ПЭТ видны: мозговая организация так называемого покоя, мозговая организация простых и все более сложных мыслительных операций и динамика этой организации, в частности, в зависимости и от того, какова направленность внимания исследуемого лица. На ПЭТ видно, какая множественная активация в мозгу происходит при восприятии и запоминании связного текста – бегущего на экране зрительного изображения (см. рис. 1). Но всегда ли это активация? На этот вопрос ответ частично уже есть, его удается находить в результатах совместных ПЭТ– и нейрофизиологических исследований.
Этот путь – от первых клинико-анатомических находок до результатов неинвазивных методов изучения нейрохимической динамики целого мозга – был пройден не без заблуждений. К одному из наиболее ярких относятся так называемые карты Клейста. По этим картам мозг, точнее его кора, была представлена в виде лоскутного одеяла, где каждый лоскуток отвечал за какую-то высшую функцию – вплоть до религиозного «я», до зон, связанных с любовью к родителям, и т. д. и т. п. Были, наоборот, забытые впоследствии драгоценные находки, в частности, «диалог» Дж. Джексона с П. Брока о зависимости речи от глубоких структур мозга (Jackson, 1885, цит. по: Jackson, 1931) или, наоборот, коры (Вгоса, 1861). Правота обоих, как это нередко бывает, выяснилась много позднее, уже в наше время.
Сегодняшний день взаимодополняющего инвазивного и неинвазивного изучения мозговой нейродинамики приоткрыл сущность того, что же кроется в зонах, высвечиваемых как активные на ПЭТ. И уже сегодня ясна целесообразность и важность сочетанного изучения мозга с помощью различных неинвазивных и при возможности инвазивных методов. При таких сочетаниях удается получить сведения о процессах в пространстве всего мозга и раскрывать физиологическую сущность этих процессов. В частности, нами было показано (Abdullaev, Bechtereva, 1993), что ПЭТ-«высвечивание» в области поля 46 (по Бродману) нижних отделов левой лобной доли (Kapur et al., 1994b; Posner, Petersen, Fox, Raichle, 1988) представляет собой дифференцированное в соответствии с фазами теста угнетение нейронной активности (торможение?), причем нейронные популяции, расположенные друг от друга на расстоянии 2 мм (разрешающая возможность метода), реагируют на разные аспекты восприятия текста – его грамматику, семантику – или, наоборот, реагируют более глобально (рис. 4).
Где же мы находимся сейчас в изучении мозговой функциональной организации мышления, как можно определить этот этап? Прошло сто тридцать шесть лет с открытия П. Брока, одного из основоположников функциональной анатомии мозга. И что же? Может создаться впечатление, что в конце XX столетия мы снова отвечаем на вопрос: где те структуры мозга, которые обеспечивают психические процессы – мышление, эмоции, память? Так в чем же принципиальная разница настоящего и прошлого в изучении функциональной анатомии?
А. Перистимульные гистограммы импульсной активности нейронных популяций 46-го поля:
1 – грамматически корректная фраза; 2 – грамматически некорректная фраза; 3 – грамматически корректная квазифраза; 4 – грамматически некорректный словоподобный набор букв
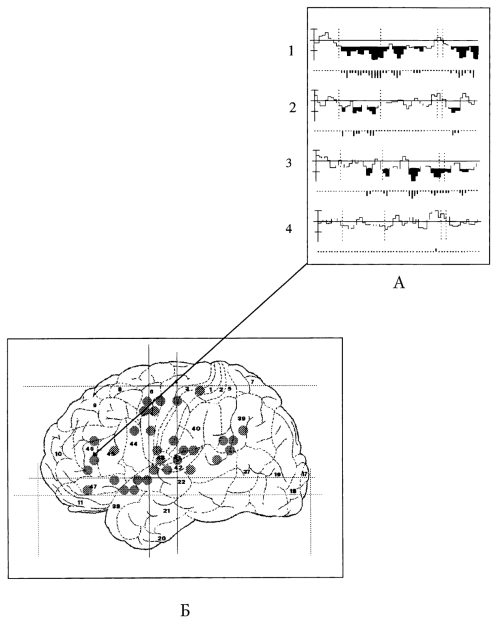
Б. Схема локализаций значимых корковых активаций при сравнении восприятия текста со счетом определенной буквы в грамматически некорректном, словоподобном наборе букв
Рис. 4. Исследование мозговой организации речи с помощью анализа импульсной активности нейронов (А) и ПЭТ (Б).
Современная техника, позволившая объявить Декаду Мозга Человека, дала возможность видеть события, происходящие в живом мозге,– их динамичность разного типа, различную у разных лиц. Мы видим сегодня не часть рисунка, а уже картину событий в мозгу, в которой прорисовываются и основной пейзаж, и некоторые детали. Если открытие П. Брока – это выявление звена системы обеспечения речи, хотя и очень важного, то сегодняшняя технология позволяет видеть множество звеньев этой системы, определять при комплексном изучении вопроса функциональную значимость этих звеньев и таким образом – иерархию системы, изучать функциональную анатомию «статистического» и индивидуального мозга человека.
Некоторые принципы и механизмы работы мозга в обеспечении мыслительной деятельности
1. Одним из наиболее общих принципов, по-видимому, следует признать сформулированный в начале XX века условно-рефлекторный (по Павлову) или сочетательно-рефлекторный (по Бехтереву) во всех его возможных вариациях. С философской точки зрения провозглашение этого принципа должно рассматриваться как существенное достижение. В изучении человеческого мозга (где исключительно велика заслуга идей и полиметодичных исследований Бехтерева) этот принцип нужно принимать во внимание при рассмотрении мозговой организации любой высшей деятельности, хотя, конечно, не следует возводить его в абсолют. Можно подчеркнуть идейную преемственность этих позиций с более ранними работами Т. Лейкока (Laycock, 1841, цит. по: Jackson, 1931) и И. М. Сеченова (1862), утверждавших рефлекторный принцип в деятельности мозга человека, и, может быть, с еще более ранними – Р. Декарта (Decartes, 1648, цит. по: Brazier, 1984).
Однако открытие рефлекторного, условно-рефлекторного (или сочетательно-рефлекторного) принципа скорее выявляет нашу общность с животным миром, чем нашу уникальность. Если рассматривать более ранние работы В. М. Бехтерева (1896), принцип развития сознания в филогенетическом ряду также, скорее, роднит весь животный мир, хотя В. М. Бехтерев подчеркивал, что местом сознательных процессов у человека являются исключительно мозговые полушария с их узлами. Более близкими науке именно о мозге человека оказываются раскрытые позднее механизмы, хотя их философское, методологическое значение существенно меньше.
2. При прямом точечном контакте с мозгом, когда записывались практически все виды физиологической активности в покое и при реализации разных видов деятельности, в том числе и мыслительной, некоторые механизмы мозга проявлялись в ходе подтверждения исходных гипотез. А на некоторые, причем весьма значимые механизмы, исследователи буквально наталкивались. Например, именно так вошел в наши знания мозговой, механизм который в научной литературе описывается как наличие у некоторых мозговых систем звеньев разной степени жесткости и который, по существу, свидетельствовал о возможности – и реальности – осуществления одной и той же деятельности не обязательно одной и той же мозговой системой, а пространственно различающимися мозговыми системами.
Этот важнейший мозговой механизм, открытый нами в 1966 году (Бехтерева, 1971; Bechtereva, 1978), далее постоянно подтверждался. Исследования с помощью ПЭТ вновь утвердили эти данные и показали, сколь существенно могут разниться мозговые системы, конечный результат деятельности которых один и тот же, исходное руководство к действию идентично 2. Сейчас к сходным представлениям пришли В. Horwitz et al. (1995), а сходные факты в своей обобщающей работе представляет Р. Е. Roland (1993). Это – один из важнейших механизмов надежности мозга, возможности правильного конечного результата мыслительной деятельности относительно независимо от внутренних и внешних помех. Разрушение (болезнь, травма) многих гибких мозговых звеньев систем организации сложной деятельности чаще всего первоначально вполне восполнимо, но постепенно лишает мозг богатства его возможностей. Очень важно для клиники, что, по крайней мере, некоторые, казалось бы, необязательные, незначимые звенья системы обеспечения, например, речевой функции, могут при необходимости взять на себя ведущую роль, определить возможность восстановления функции при необратимой гибели главного звена соответствующей системы, в частности, зоны Брока.
В обеспечении разных видов деятельности, и в том числе мыслительной, мозг обладает еще целым рядом механизмов надежности, увеличения его возможностей. Речь здесь идет о явной или латентной полифункциональности очень многих нейронных популяций, которая может присутствовать исходно (явная) или проявляться при изменении химических модуляционных влияний (латентная: Бехтерева, 1971; 1980). И наконец, не ставя перед собой здесь задачу перечисления всех механизмов надежности работы мозга, упомянем, как привлекший сейчас большое внимание, механизм детекции ошибок.
Впервые феномен детекции ошибок, «детектор ошибок», был открыт нами в 1968 году (Bechtereva, Gretchin, 1968). С тех пор различные аспекты вопроса рассматривались нами в большом числе публикаций и в главах ряда монографий (Бехтерева, 1971; 1974; 1980; 1988а; 1990; 1994а; Bechtereva, 1978; 1987; Bechtereva, 1984; Bechtereva, Kropotov, 1986; Bechtereva et al., 1990a; 1991). Было показано, что в мозгу имеются нейронные популяции, на какую-то данную сложную деятельность не реагирующие; реагирующие на ее правильное выполнение; реагирующие и на правильное, и на ошибочное выполнение задания. И наконец, отдельные нейронные популяции реагируют именно при ошибочном выполнении деятельности, будь то в связи с дефектом восприятия (ранняя реакция) или дефектом реализации (поздняя реакция).
Такие нейронные популяции были обнаружены нами первоначально в подкорковых структурах. Позднее такие же нейронные популяции были обнаружены нами и в коре.
Детектор ошибок активизируется при рассогласовании деятельности с ее планом, точнее, с хранящейся в мозгу матрицей. (Понятно, что вряд ли он активируется при ошибках в деятельности творческой.)
3. Детектор ошибок был заново «открыт» при некоторой вариации нейрофизиологической методики (вызванные потенциалы, а не динамика нейронной активности) рядом исследователей, причем был обозначен совершенно так же, как детектор ошибок (Gehring et al., 1993; Dehaene et al., 1994; Bernstein et al., 1995). Несколько ранее принципиально то же явление было описано (и приобрело очень широкое звучание) R. Naatanen (1992) и обозначено, как Missmatch Negativity. По существу, речь идет о рассогласовании с планом, появлении неожиданного элемента для матрицы-схемы ситуации или действия. Феномен этот благодаря энтузиазму R. Naatanen оказался более изучен. И кроме того, еще и потому, что являлся основной задачей его лаборатории, а в нашем случае – лишь интересной находкой на пути широкого изучения нейрофизиологических механизмов психики.
Нейрофизиологическое исследование мозговой организации мыслительной деятельности обнаружило множество интересных механизмов и свойств в этой работе мозга. Так, в динамике сверхмедленных процессов проявились защитные механизмы мозга. Они характеризовались разнонаправленными сдвигами этих базисных физиологических показателей и особенно значимы как превентивный механизм, препятствующий распространению в мозгу эмоционально-детерминированных сдвигов сверхмедленных физиологических процессов, в свою очередь определяющих нежелательное развитие патологических эмоций. Отдельно нужно рассматривать изменения в ЭЭГ, отражающие защитный механизм другого типа, способствующий торможению избыточной информации. Наиболее ярко это проявляется при эпилепсии, где развитие заболевания отражает в том числе и недостаточность защитных механизмов мозга, а по мере этого развития оказывается возможным наблюдать увеличение амплитуды и длительности пароксизмальных проявлений на ЭЭГ (а также увеличение продолжительности ее элементарных составляющих). Как и любой защитный процесс организма, при своем усилении этот механизм легко становится патологическим (Бехтерева, 1988а; 1990).
Тема механизмов мозга поистине неисчерпаема. Здесь, пожалуй, целесообразно для стимулирования интереса к вопросу привести лишь еще один тип наших наблюдений, как и многое в сложной проблеме, может быть, и нуждающийся в дальнейшем изучении. Речь идет о пространственно преимущественно тормозных реакциях коры и преимущественно активационных реакциях подкорки при активации в коре лишь зон, имеющих первостепенное значение именно для данной деятельности. Такого рода соотношение наблюдалось нами в ходе реализации различных психологических проб (Bechtereva et al., 1990b). Это проявлялось по миновании периода первоначальной генерализованной активации как ориентировочной реакции, являющейся одним из главных механизмов самосохранения мозга (Bechtereva et al., 1972). Если приведенные данные будут подтверждаться (похожие сведения приводятся в работах: Hoshi et al., 1994; Schwartz et al., 1996), придется, может быть, пересматривать многие из уже сложившихся представлений о корково-подкорковых соотношениях в обеспечении мыслительной деятельности.






