Импульсная активность нейронных популяций при мыслительной деятельности
Первый этап исследования. Мы имели возможность находить точки мозга, более или менее тесно связанные с психической деятельностью. Для расшифровки протекающих в них процессов физиологу предстояла сначала задача выбрать из регистрируемых биоэлектрических процессов – наиболее подходящий для этой цели. Первым (хотя и не единственным) кандидатом на данную роль была импульсная активность нейронов, прежде всего, в связи с быстродействием мыслительных процессов. Далее встал вопрос: отдельная нервная клетка или функционально объединенное сообщество нервных клеток? Многолетний опыт экспериментальных исследований (Василевский, 1968; Альтман, 1972, и др.) показал, что одна нервная клетка чаще всего не способна отражать своей активностью сложные сигналы. При регистрации динамики отдельной или, точнее, отдельных клеток как представительных единиц в человеческом мозгу можно получить ряд интересных сведений о его работе (см. выше). Однако на уровне группы нервных клеток, работающих в том нужном согласии друг с другом, которое создает новое качество, можно было надеяться найти отражение уже более сложных сигналов (Коган, 1973). Вопросы о свойствах популяции нервных клеток подробно рассматриваются Буллоком (Bullok, 1979).
Таким образом, еще в период, когда сравнительно редко использовалось отведение импульсной активности не с микроэлектродов, а с так называемых микро-макроэлектродов (в нашем случае с диаметром 50–100 мкм и рабочей поверхностью от 0.01 до 0.15 мм2), было начато изучение реорганизации импульсной активности нейронных ансамблей при простейших видах мыслительной деятельности. С вживленных в различные структуры мозга (кору, образования таламуса и стриопаллидарной системы, верхние отделы ствола, медиобазальные отделы височных долей) электродов регистрировалась активность популяций нервных клеток. В этой активности обычно обнаруживались пики разрядов нескольких нервных клеток, более или менее существенно превышавшие по амплитуде труднодифференцируемую активность большого количества нервных клеток. Варианты регистрируемой с вживленных электродов активности представлены на рис. 14.
Любое исследование тонких нейрофизиологических коррелятов мыслительной деятельности до последнего времени имело и еще, по-видимому, в течение ряда лет будет иметь, кроме собственно нейрофизиологических, по крайней мере два аспекта. Так, во-первых, в каждом из них в соответствии с нейрофизиологическими задачами требовалось найти адекватный психологический подход. И хотя многие из применявшихся тестов строились на основе классических психологических проб, далеко не все могло быть использовано в традиционном варианте (тест Вине и др.). Во-вторых, возможности нейрофизиологического анализа ответов активной, реагирующей на психологический тест точки существенно зависели от степени совершенства математико-технической обработки извлечения полезного сигнала.

Рис. 14. Примеры нейронограммы мультиклеточной активности различных зон коры и подкорковых структур головного мозга человека.
Сверху вниз:
поясная извилина, премоторная область коры, вентролатеральное таламическое ядро срединный центр, заднее вентральное латеральное таламическое ядро, бледный шар. Калибровка: 20 мс, 50 мкВ
Для избежания разночтений уточним, в чем разница между популяцией нервных клеток и их рабочим ансамблем. С электрода регистрируется активность группы нервных клеток. Это – всегда популяция, но не всегда ансамбль. Как можно проверить подобное утверждение? Дается задача, одна, другая – сосчитать что-то в уме, повторить ряд слов. Затем с помощью специальных программ и ЭВМ изучается реакция этой популяции нервных клеток. Какие здесь возможны варианты? Может просто не произойти ничего существенного – и тогда, скорее всего, данная популяция не имеет отношения к реализуемой деятельности. Реакция может быть относительно простой – в виде общего тонического увеличения или уменьшения частоты разрядов. В этом случае следует предположить, что данная популяция вносит в систему обеспечения психической деятельности «энергетический» вклад того или другого знака или что существующие методы еще не способны обнаружить более тонкие изменения. Может выявиться, наоборот, перестройка организации работы нервных клеток. И тогда правомерно использовать термин «ансамбль», хотя в конкретном случае это может быть лишь часть ансамбля, так как остальная находится за пределами возможностей регистрации активности ее клеток с данного электрода. А может быть и иначе. Только часть популяции существенно реорганизуется, работает в данных условиях как слаженный ансамбль. Но вот задается другая деятельность, ансамбль захватывает другие клетки из популяции или, наоборот, становится меньше. Ансамбли очень динамичны, они все время «дышат». Наверное, тем, кто не работает непосредственно в данной области, представить себе организацию и реорганизацию ансамблей можно, вспомнив динамичную световую рекламу, – из набора однотипных источников света слагаются ежемоментно меняющиеся, иногда очень сложные световые узоры. Понятно, что не только это сравнение, но и приведенные выше положения – лишь схема, которую иногда очень нелегко уточнить в процессе математико-технической обработки.
Те зоны, в которых нами обнаруживались воспроизводимые изменения физиологических показателей мозга при психологических пробах, не относились к анатомическим образованиям акустического (слухового) анализатора. Лишь теперь, через много лет после начала наших исследований, у тех, кто работает в области мозгового анализа звуковых сигналов, начал пробуждаться интерес именно к этим зонам, в которых, как предполагают Эммере (Emmers, 1970) и Я. А. Альтман с соавторами (1979), происходит обработка сложных сигналов, высшая по отношению к протекающей в классических образованиях анализатора. И, как считает Б. Ф. Толкунов (1978), именно в этих зонах существуют оптимальные анатомо-физиологические условия для высшей интеграции. Тем не менее самым началом нашего углубленного изучения тонких нейрофизиологических коррелятов психической деятельности были поиск и отражения акустических характеристик слов и их элементов в активности популяций (ансамблей) нейронов структур стриопаллидарной системы, таламуса, медиобазальных отделов височных долей, верхних отделов ретикулярной формации не как дань представлениям Пенфильда (Penfield, 1975) о центроэнцефальной системе, а как результат собственного опыта. Как указывалось, именно в этих зонах отмечались воспроизводимые реакции при психологических пробах. При этом мы отдавали себе отчет в том, что поиск коррелятов может быть затруднен непрямым отражением акустических свойств сигнала в связи с характером образований.
В результате первых исследований, по ходу которых проводилась весьма трудоемкая отработка методов анализа мультиклеточной активности, была показана, хотя и непрямая, связь развивающихся в области этих подкорковых структур пространственно-временны?х перестроек импульсной ак тивности с частотно-амплитудными характеристиками входного сложного звукового сигнала – слова (Бехтерева и др., 1971). Известную роль в прогрессе исследований сыграло предложенное на основе работ Сондерсона с соавторами (Sanderson et al., 1973) и П. В. Бундзена (1976) использование так называемой дистрибутивной формы представления пространственно-временны?х перестроек импульсной активности нейронных популяций. Эта форма не только открыла новые возможности машинного анализа импульсной активности при психологических пробах, но и явилась необходимым методическим толчком для изучения значимых элементов в перестройках мультиклеточной активности. Детали математико-технической обработки импульсной активности приведены в работах П. В. Бундзена с соавторами (1975б), Ю. Л. Гоголицына, П. Д. Перепелкина (1975), Ю. Д. Кропотова (1976, 1978), П. Д. Перепелкина (1977, 1978), в разделе П. Д. Перепелкина в работе Н. П. Бехтеревой с соавторами (1977а). Идет постоянное совершенствование методов на основе общей принципиальной схемы анализа. По ходу изложения материалов там, где без этого нельзя обойтись, будет приведена физиологическая логика этого математико-технического анализа.
Как видно из наших первых работ, слово как сложный акустический сигнал и – что в этих работах не исключается, но и не доказывается, – как сигнал семантически значимый вызывает тонические и фазические перестройки импульсной активности. Одновременно в данных работах показано, что упомянутые перестройки обнаруживаются только в части популяций, а ансамблем может являться часть большой популяции. Как это обнаруживается? Если при работе с отдельными нервными клетками для получения представления о зоне мозга исследователи идут неизбежно по пути накопления данных о поведении нейронов в этой зоне, то при работе с популяциями нервных клеток методической основой последующей интегративной фазы, объединения, является первоначальное разделение популяций на достаточно малые группы нейронов. Осуществляется выделение тех разрядов нейронов, которые, превышая уровень физиологического шума, позволяли проводить исследования при минимально возможных искажениях мозговых сигналов – имеется в виду прежде всего искажение формы разряда при регистрации в результате суперпозиции двух и более спайков. С этой целью мы прежде всего разделяли разряды по амплитуде. Такой метод при некоторой его уязвимости и используется не только у нас, но и в ряде других исследований (Keidel, 1975; Schwartz et al., 1976; Halgren et al., 1978; Ravagnati et al., 1979). Разделение по амплитуде производится в виде так называемой «оконной» дискриминации (Бехтерева и др., 1973), предполагающей выделение нескольких амплитудных уровней разрядов. При этом в каждое «окно» попадают именно те разряды, пик которых оканчивается в его пределах. Именно на основе этой оконной дискриминации и были разделены разряды мультиклеточной активности в наших первых исследованиях.
Наравне с «оконной» дискриминацией в связи с накоплением достаточного материала и отсюда возможности выбора сейчас предпочтительно используется определенный амплитудный порог, зависящий от соотношения амплитуд выделяемых отдельных разрядов и физиологического шума более низкой амплитуды, «щетки», искажающих друг друга разрядов многих клеток; обычно этот порог определяется в размере 0.7 от максимальной амплитуды разрядов. При этом, естественно, учитывается лишь часть регистрируемой активности, но, что очень важно, ее наименее искаженная или даже практически неискаженная часть.
В обоих случаях обычно оказывается возможным обнаружить большее или меньшее количество разрядов равной и разной амплитуды. При исследовании активности отдельных нейронов амплитудный показатель может до известной степени и на небольшом промежутке времени помочь идентифицировать, опознать нейрон. Сходно обстоит дело и при дифференциации мультиклеточной активности. Однако идентификация отдельных нейронов более надежна, хотя тоже не абсолютна, при использовании и амплитудного, и дополнительного показателя – формы потенциалов действия. Этот прием также с 1973 года в течение ряда лет использовался в нашей лаборатории.
При регистрации мультиклеточной активности оказывается возможным выделить с небольшими искажениями динамику активности до четырех, редко более, отдельных нейронов, то есть принципиально то же, что возможно и в условиях аналогичного собственно микроэлектродного исследования.
Существенный выигрыш за счет регистрации именно мультиклеточной активности, а не активности отдельных нейронов, связан с возможностью изучения в таких условиях взаимодействия не только удаленных нейронов, но и нейронов, расположенных заведомо в пределах рабочего ансамбля. В эксперименте этот вопрос мог бы быть решенным за счет введения ряда микроэлектродов с заданным и фиксированным расположением между ними, однако до последнего времени такой прием в клинических исследованиях является неоправданным, несмотря на работы в области зрительного и слухового протезирования (Simmons et al., 1965; Brindley, Lewin, 1968; Brindley, 1970; Michelson, 1971). He исключено также, что для нейрофизиологического анализа это не представит особых преимуществ, хотя данный вопрос лучше оставить пока открытым. Проблема чистоты получаемых данных решается нами иначе – применением в исследованиях мозга человека электродов, у которых возможно произвольно варьировать рабочую поверхность и, таким образом, одновременно или последовательно записывать большее или меньшее количество нейронов (Матвеев, Киселев, 1978). Взаимодействие нейронов в пределах одной зоны мозга и между разными его зонами определялось первоначально путем сравнения активности, ее текущей частоты предварительно дискриминированных (способом оконного различения и др.) нейронов, причем основным приемом служил факторный анализ, а эпоха анализа избиралась в соответствии с задачами и плотностью выделенной активности.
Далее с этой целью применялись и другие приемы, и прежде всего изучение регулярности и связности импульсного потока в форме разных видов интервального анализа и прием когерентной детекции – выявления синхронности событий в удаленных зонах мозга (Бундзен и др., 1975б; Кропотов, 1976, 1978; Перепелкин, 1977; Бундзен, Давид, 1979). Эти исследования проводились с целью уточнения не только взаимодействия между нервными элементами, но и всех перестроек местной активности нейронных популяций, характерных для исследуемого вида деятельности. Стимулирующими факторами данного направления работ служили следующие.
1. Экспериментальные исследования, в которых описывалась регулярность и связность импульсных потоков в активности отдельных нейронов и нейронных популяций и возможность развития микроструктурных особенностей паттернов межимпульсных интервалов – узоров межимпульсных разрядов (Gerstein, 1960; Poggio, Vernstein, 1964; Kiang, 1965; Nakahama, 1966; Perkel et al., 1967;. Perkel, Bullok, 1968; Segundo, Perkel, 1969; Verzeano et al., 1970; Василевский и др., 1972; Babb et al., 1973; Василевский, 1976; Nakahama et al., 1977; Толкунов, 1978; Чораян, 1978; Шульгина, 1978, и многие другие).
2. Специальное изучение чисто частотного кодирования, проведенное Эхорном с соавторами (Echorn et al., 1976), показавшее, что при этом виде кодирования возможные потери информации могут быть очень велики – от 10 до 95 %!
3. Данные о функциональных и морфологических основах возможности кодирования появлением специфических узоров импульсной активности (Anderson et al., 1964; Schlag, Villabanca, 1968; Gerstein, Perkel, 1969; Emmers, 1970, 1976; Moore et al., 1970; Sanderson et al., 1973; Katayama, Murata, 1974; Baldissera, Parmiggiani, 1976; Толкунов, 1978).
4. Известная закономерность об усилении роли кодов в процессе эволюционного развития и становления высших форм деятельности (Ашмарин, 1975; Николаев, 1976; косвенно данные: Богданов, 1978).
Уже в самом начале изучения взаимодействия нейронов было обнаружено, что в процессе психической деятельности наряду с изменением текущей частоты разрядов происходят и другие характерные изменения импульсной активности нейронных популяций. Ее местная перестройка в исследуемых структурах при психической деятельности характеризовалась не только изменением частоты, но и появлением удерживающейся в течение секунд, десятков секунд и – реже – минут мозаики и динамики пространственных взаимоотношений между близлежащими и удаленными нейронами в дистантно расположенных зонах мозга, что выражалось, в частности, появлением определенной структуры разрядов нейронной популяции – их определенного паттерна.
Именно поэтому в физиологическом плане вначале для, скажем так, проверки «проходимости пути» выгодной оказалась дистрибутивная форма представления развивающейся при исследуемой деятельности перестройки паттерна, так как она сохраняла информацию о пространственно-временны?х отношениях нейронов и оставляла возможность их последующего, более углубленного исследования.
Дистрибутивная форма представления паттерна отражала динамику межимпульсных интервалов между разрядами группы нейронов, причем количество исследуемых (записываемых) нейронов определялось уровнем амплитудного среза. В плане возможностей исследования мультиклеточной активности с помощью ЭВМ при применении эталонного поиска паттерн в дистрибутивной форме в виде конечной (ограниченной временем) численной последовательности длительностей интервалов представлял собой приемлемый эталон, который мог быть заложен в ЭВМ (рис. 15) (Перепелкин, 1977). У каждого больного для каждой нейронной популяции в определенные отрезки времени накапливались дистрибутивные представления паттернов мультиклеточной активности при восприятии, удержании в памяти и воспроизведении различных слов (Бундзен и др., 1975б). Так формировался банк эталонов, позволяющий у того же больного в соответствующих нейронных популяциях и обычно в связи с кратковременностью жизни этих паттернов в той же записи обнаруживать момент появления соответствующих паттернов и таким образом определять возникновение «слов» в мозгу не только в соответствии с непосредственным содержанием входного и выходного сигналов – предъявляемых слов или изображений и ответно произносимых слов, но и того, что имело к этим сигналам смысловое косвенное отношение. Следует подчеркнуть, что в связи с указанными выше временны?ми ограничениями работы с эталонным поиском целесообразнее всего проводить (что и проводилось далее) в реальном времени исследования.
Вслед за использованием дистрибутивного представления паттерна начался поиск его значимых элементарных составляющих.
Рис. 15. Реальная последовательность межимпульсных интервалов, выявленная путем анализа мультиклеточной активности (срединный центр) при воспроизведении испытуемым всех отдельных цифр (фото с дисплея анализатора импульсов «Didac-4000»).
Размерность каждого прямоугольника намеченной сетки – 10 мс (по вертикали), 10 интервалов (по горизонтали). По оси ординат – значение межимпульсных интервалов, мс; по оси абсцисс – последовательность межимпульсных интервалов по порядку. Подчеркнуты моменты воспроизведения различных цифр
В основу был положен тот же принцип – обнаружение связностей, групповых последовательностей разрядов с определенными соотношениями интервалов. Важность задачи поиска элементарных составляющих пространственно-временно?й реорганизации мультиклеточной активности, развивающейся при восприятии, удержании в памяти и воспроизведении слов, определялась рядом положений. Прежде всего дистрибутивное представление паттерна, полученное на основе оконной дискриминации или амплитудного среза, не обязательно содержало только полезный сигнал. Возможность использования паттерна в эталонном поиске для обнаружения с помощью ЭВМ аналогов в мультиклеточной активности показывала значимость его как выражения тонких нейрофизиологических коррелятов слов. Но ни этот прием, ни какой-либо другой из применявшихся ранее, подтверждающие значимость фактора длительности межимпульсных интервалов в паттерне, не свидетельствовали о чистоте данного представления паттерна, не давали сведений о соотношении в нем сигнала и шума.
Эталонный поиск использовался в физиологии и ранее. Так, например, без особой необходимости для диагностики, но как важный элемент последующего управления биоэлектрической активностью больных эпилепсией эталонный поиск с помощью ЭВМ применялся уже в 60-х годах при использовании в качестве эталона эпилептиформной активности комплекса пик—волна. В этом случае, как вполне понятно, феномен спайк—волна мог быть выделен и при использовании менее информативного эталона – только спайка или только медленной волны. Но тогда наравне с сочетанным биоэлектрическим феноменом на ЭЭГ неизменно выделялись бы (распознавались!) и отдельные, и групповые спайки, и различные медленные волны. Использование дистрибутивного представления паттерна мультиклеточной активности принципиально могло быть таким же. Дистрибутивный эталон мог также представлять собой как бы часть сигнала. Образно (а может быть, и не очень образно, а реально?) он отражал бы часть нейрофизиологических коррелятов слова – его слог, триграмму, встречающиеся и в других словах. С другой стороны, в дистрибутивном представлении в зависимости от ряда условий и прежде всего просто от уровня амплитудного среза импульсной активности может содержаться и избыточная информация, загрязняющая эталон и, как это ни парадоксально, другим путем ведущая к тому же результату – возможности опознания с его помощью не только тех лов, на основе нейрофизиологического отражения которых он был образован.
Сложность вопроса усугублялась тем, что паттерн мог обнаруживаться в мозгу и в полном, развернутом виде, по времени соответствующем слышимому или произносимому слову, и – особенно фазу удержания слов в памяти или считывания из памяти – в сжатой, компрессированной форме. Действительно, уже в самых первых исследованиях в нашей лаборатории было показано, что после восприятия слова непосредственно за развернутым, полным паттерном в мозгу появляется компрессированный паттерн, содержащий опорные элементы первого.
Этим не кончаются сложности (хотя их не следует и переоценивать!) работы с «неочищенным» паттерном и любой формой его представления. Так, например, при эталонном поиске наиболее адекватным оказалось использование дистрибутивного представления не только той перестройки импульсной активности, которая соответствует по времени моменту произнесения слова больным, но и несколько большего ее отрезка, увеличенного за счет перестройки, предшествующей ответу, за счет захвата в эталон и фазы формирования ответа. Значимость этой фазы для ответного речевого сигнала подтверждается в ряде работ и прямо, в частности, в исследованиях С. Н. Раевой (1972, 1977), Хелгрина с соавторами (Halgren et al., 1978), и косвенно – в работах другого направления, связанных с регистрацией состояний мышц при произнесении слов (Syka, Schramm, 1975).
Все эти и ряд других соображений определили работу по выделению более элементарных составляющих паттерна.
Если первоначально за основу был взят паттерн, соотносимый со словом или его элементом, то далее методически и технически задача решалась самыми разными способами, в том числе изучением фоновой активности и активности при выполнении психологических тестов с соотнесением данных с фоновыми и тестовыми не только a priori, но и a posteriori проведенной обработки.
Математико-технический аспект данных направлений исследования мультиклеточной активности был описан в 1977 году. Здесь важно подчеркнуть, что в результате проведенных работ (Бехтерева, Бундзен, 1974б; Гоголицын, 1974; Перепелкин, 1974; Бундзен, 1976, 1977; Малышев, 1977; Шкурина, 1978) в отрезках записи мультиклеточной активности продолжительностью обычно в несколько десятков секунд были выделены групповые последовательности разрядов с фиксированным ( +10 %) интервалами, причем функционально значимым элементом общего паттерна оказывались последовательности из трех и более интервалов. При этом, хотя и наблюдались групповые последовательности разрядов с определенными интервалами в количестве более 3 (до 6–7), они были менее стабильны, уникальны и менее воспроизводимы. Это позволило считать последовательность из трех интервалов минимальной и достаточной для физиологической роли ее как элемента паттерна. Чем характеризовалась функциональная значимость групповой последовательности разрядов с определенными интервалами? Первым опорным критерием было появление или существенное учащение этих триад в период психологического теста, соотносимое с его фазами, воспроизводимое у одного того же больного в аналогичных условиях. Вторым критерием была возможность использования групповой последовательности разрядов в качестве образца при эталонном поиске. Иными словами, если использование какой-то данной триады в качестве эталона позволяло выявлять в мультиклеточной активности моменты появления в мозгу общих паттернов, в которых присутствовала данная триада, она расценивалась как значимый элемент паттерна. Не перебирая все критерии, следует подчеркнуть еще один, не относящийся к конкретной триаде или квадриаде и т. п., но характеризующий значение этих групповых последовательностей как элементов паттерна, являющегося тонким нейрофизиологическим коррелятом психических процессов. Это – показанная В. Н. Малышевым (1979) возможность встраивания триад в паттерн при введении квазислова в смысловое поле определенных слов. Естественно, при этом удовлетворялись и первые два критерия.
Что представляет собой фоновая, или, точнее, так называемая фоновая, активность с точки зрения возможности определения в ней групповых последовательностей разрядов с определенными интервалами? Можно ли обнаружить в фоновой активности те же триады? Нередко можно, но обычно с меньшей вероятностью (Перепелкин, 1978). Что действительно обнаруживается в фоне в большом количестве, так это парные интервалы с определенной их длительностью, в том числе и одинаковой в обоих интервалах. Именно парные интервальные последовательности, по-видимому, служат физиологической основой образований триад (Малышев, 1977), а методически являются основой одного из наиболее перспективных приемов их поиска (Бундзен, 1977). Обнаружив парные интервалы, дальше с помощью соответствующих программ мы исследовали, не «привязывается» ли к ним в связи с тестом и не удерживается ли до или после этой пары еще один интервал, образуя таким образом триаду. По-видимому, парные интервалы могут рассматриваться в качестве опорных жестких единств для формирования интервальных триад. Однако для понимания механизмов формирования значимых групповых последовательностей разрядов с тремя и более интервалами определенной продолжительности нуждается в дальнейшем уточнении вопрос о парных интервальных последовательностях – являются ли они результатом развертывающихся генетических свойств мозга, процесса обучения в общем виде или обучения более конкретного, речевого.
Одна триада на определенном отрезке времени исследования может быть функциональным элементом нейрофизиологического паттерна и служить эталоном для машинного исследования импульсной активности одного или нескольких близких по смыслу слов. Однако это не общее правило, единственной триадой паттерн не исчерпывается даже в одной нейронной популяции. Каждый паттерн состоит из совокупности групповых последовательностей разрядов, и, таким образом, отражение слова или его элемента в импульсной активности нейронных популяций характеризуется набором этих групповых последовательностей разрядов. Говоря об интервальных последовательностях, следует помнить, что функционально значимым фактором является пространственно-временна?я перестройка межнейронных отношений, а они – ее отражением. При этом в разных, близких и удаленных друг от друга, зонах мозга одно и то же слово или его элемент, как правило, вызывает появление паттернов, состоящих из групповых последовательностей разрядов, главным образом своих для каждой зоны мозга, но также и очень сходных для разных зон (Бехтерева и др., 1977а). В 1980 году нами высказывалось предположение, что сходные групповые последовательности разрядов являются своего рода объединяющим, системообразующим фактором. Эти предположения нашли прямое подтверждение в исследованиях Г. А. Вартаняна и А. А. Пирогова (1987). Что же касается разных групповых последовательностей разрядов в разных зонах мозга, то они, повидимому, характеризуют прежде всего распределенность паттерна – распределенность кодирования психических процессов (Бехтерева, 1977). Исследование общих элементарных единиц и синхронных разрядов в разных нейронных популяциях, а также в разных элементарных единицах является задачей дальнейших исследований. Предполагается, что результаты этих исследований могут пролить свет на вопрос о том, какое новое качество приобретает популяция по сравнению с отдельной нервной клеткой, несколько звеньев системы – по сравнению с одним звеном, система – по сравнению со звеном или совокупностью звеньев. Надо подчеркнуть, что этот вопрос ставился нами с первых шагов данного исследования (Бехтерева, 1966, 1971).
При прослеживании динамики синхронных разрядов в работающих при тесте на принятие решения нейронных популяциях видно, что процесс принятия решения характеризуется динамическим увеличением синхронизации – сходства в разных зонах мозга. Есть известные основания полагать, что дальнейшее изучение проблемы приведет к обнаружению в паттернах и на уровне системы, и на уровне нейронных популяций жестких и гибких элементов (более или менее устойчивых). Это не противоречит основному положению о распределенности перестроек, но подтверждает общность принципа жестких и гибких элементов на разных уровнях организации мозга.
Чем дольше продолжалось изучение вопроса, тем больше подтверждалось, что ансамбль по сравнению с суммой клеток обладает дополнительными свойствами. При исследовании амплитуды и формы спайков оказалось, что групповые последовательности разрядов с определенными интервалами, и в частности триады, образуются (могут образовываться!) за счет разных нейронов, а не одного и того же нейрона. Таким образом, принцип пространственно-временно?й организации перестроек импульсной активности обнаружился и на уровне элемента паттерна.
На сегодня это можно объяснить с помощью следующей гипотезы: ансамблем нейронов является функциональное единство из ряда групп нейронов, каждая из которых обладает своими свойствами. При изменении состояния в активной зоне мозга под влиянием тех или иных факторов (приходящий к нейронам сигнал, изменение уровня базисных медленных электрических процессов и т. д.) срабатывает один нейрон из группы сходных нейронов, причем другие сходные нейроны в этом случае тормозятся, а нейрон из другой группы разряжается через временной интервал за счет прежде всего синаптической задержки при не всегда прямой передаче возбуждения (разного рода прямые и обратные связи). При этом синаптическая задержка играет роль квантующего фактора. Естественно, возможны и другие, также гипотетичные, варианты, в частности те, которые обсуждаются в работах Эммерса (Emmers, 1970) и были упомянуты нами выше. Морфологические данные (Поляков, 1973) о наличии разных нейронов и глиальных клеток даже в небольших клеточных скоплениях мозга как будто бы не противоречат такой точке зрения. Но, естественно, этот вопрос нуждается в дальнейшем изучении. Вопрос все тот же – часть и целое, нейрон и ансамбль и т. д.
При обсуждении методологических проблем современной нейрофизиологии уже говорилось о том, что далеко не все применяемые в психологии методы могут быть использованы в комплексном исследовании при изучении нейрофизиологических коррелятов психической деятельности. Психологический аспект исследования должен соответствовать нейрофизиологическим задачам. При этом на разных этапах исследования требования к психологической его части различны.
На том этапе, когда задачей нейрофизиологического исследования становится выделение первоначально общих перестроек импульсной активности в связи с конкретными словами, а затем все более тонких элементов этих перестроек, тесты существенно меняются. Так, например, произносятся одиночные синтетические гласные фонемы – ответ больного заключается в воспроизведении этой фонемы или того, что по его мнению, было ему предъявлено (случай предъявления так называемой смешанной синтетической фонемы, которая могла бы быть распознана и как [А], и как [Е]). Произносятся слоги или триграммы, причем их немного, нередко – один слог или одна триграмма, больной повторяет их в зависимости от задания сразу же или по команде. Называются отдельные слова или маленькие группы слов – знакомых, часто или редко встречающихся в речи, квазислова, слова на незнакомых больному иностранных языках; больной повторяет их также непосредственно после предъявления или по команде. Все это достаточно просто для больного, и какая-либо собственно психологическая сложность задачи при этом отсутствует. Однако, как вполне понятно, именно этот этап исследования исключительно важен не только для нейрофизиологии, но и для психологии как науки. Только при сочетанном психонейрофизиологическом исследовании может быть получен надежный ответ на вопрос, что реально происходит в мозгу при восприятии слова, в момент, когда формируется оперативная единица психической деятельности, в момент подготовки и реализации вербального ответа и т. д. Существующие представления о наличии в мозгу возникающей в процессе обучения «фонемной решетки» (Жинкин, 1958) интересны, но, вероятно, не могут претендовать на абсолютную универсальность.
Рабочей гипотезой в проводимых исследованиях были представления о том, что слово может быть отражено в мозгу пофонемно, в виде отдельных слогов и больших блоков, особенно при частом употреблении его в речи. При этом принцип пофонемного кодирования остается, по-видимому, неизменным при обучении и вербализации ответа. Однако, чтобы использовать получаемые нейрофизиологические данные для понимания механизмов собственно мыслительной деятельности, необходимыми этапами были исследование отражения в выделяемом импульсном паттерне смысловых свойств слова и далее построение и предъявление задач, позволяющих судить о нейрофизиологических основах собственно мыслительной деятельности человека. Зоны, участвующие в обеспечении психической деятельности, достаточно достоверно можно было обнаружить вначале с помощью метода постстимульных гистограмм (Гоголицын, 1979; Кропотов, 1976б), а в последующие годы – перистимульных гистограмм (ПСГ) (Гоголицын и др., 1987а).
В этом случае удается обнаружить (например, в передних отделах поясной извилины) и некоторую разницу реакций на слова разных смысловых полей (рис. 16). Однако вполне понятно, что прямое решение этой задачи если не невозможно, то во всяком случае весьма сложно, а поэтому изучение отражения смысловых свойств в перестройках импульсной активности нейронных популяций было предпринято сотрудниками нашей лаборатории в форме поисков признаков смысловой общности или, что в данном случае принципиально то же самое, смысловой дифференциации на основе модификации теста Озгуда (Osgood, 1953).
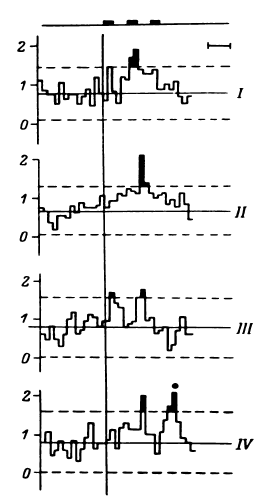
Рис. 16. ПСГ активности нейронной популяции переднего отдела поясной извилины.
Предположительный объем популяции – 2–3 нейрона. Каждая гистограмма накоплена по 16 предъявлениям пар слов, которые обобщались по смыслу пациентом. По оси абсцисс – время (калибровка – 1 с, бин гистограммы – 200 мс). Зачерненные прямоугольники вверху – средние эпохи звучания двух предъявляемых слов и ответа пациента. Вертикальная линия – начало теста. По оси ординат – среднее число разрядов нейронов в бине гистограммы за одно предъявление. Штриховыми линиями ограничен доверительный интервал, соответствующий 99%-ной вероятности отклонения частоты импульсации от уровня фона. I – гистограмма, накопленная при предъявлении пар слов, обозначающих предметы мебели; II – то же – посуды; III – то же – названий деревьев; IV – смешанная группа (пары слов не могли быть обобщены и пациент отвечал «нет»). Черный кружок – дополнительный всплеск активности, возможно, эмоциогенный. Заметно также большее возрастание частоты разрядов при предъявлении второго слова
Больному назывались последовательно слова, которые могли бы быть обобщены по смыслу каким-то другим понятием. Так, например, перечислялись предметы мебели: «стул», «стол», «шкаф» и т. д., после чего больного просили назвать обобщающее слово, как вполне понятно, в данном случае – «мебель». Задача могла быть поставлена и в еще более облегченной для больного форме – ее условия формулировались заранее: «Вам будет назван ряд слов. Постарайтесь уловить в них общий смысл и назвать обобщающее понятие». Если в тестах перечислялись названия цветов, больной отвечал: «Цветы», если деревьев, – ответ был: «Деревья».
Далее предпринималась задача встраивания нового слова в смысловое поле, сформировавшееся у больного в онтогенезе на основе биосоциального опыта (Малышев, 1979). Так, например, попеременно перечислялись названия цветов и деревьев и в ходе теста (обычно не в начале и не в конце) без предупреждения о том, что в смысловом поле будет предъявлен не имеющий смысла элемент, произносилось квазислово. Если больной спрашивал, что это такое, в ходе теста ему отвечали, что это – название цветка или дерева. В последующем больному сообщалось, что данный словоподобный сигнал значения не имеет. При изучении нейрофизиологического отражения смысла слов применялись пробы, где одно и то же по звучанию слово имело в зависимости от контекста разный смысл. Так, например, смысл слова «коса» оказывался различным, если в задании назывались «русая коса» или «острая коса». Так же направленно менялось значение слова «ключ» («ключ к замку», «лесной ключ») и т. д.
Результаты исследований нейрофизиологического выражения смысловой общности использовались далее для изучения нейрофизиологического выражения простейших мыслительных операций. Мыслительные операции, нейрофизиологическое выражение которых исследовалось нами, являлись несложными для любого человека. Однако исследование строилось таким образом, что его результаты могли служить не только непосредственному психонейрофизиологическому исследованию, но и переходу к более сложным задачам. Предъявлялись цветные изображения, задавался вопрос, ответить на который было сравнительно легко на основе простейшего анализа компонентов изображения.
Например, на слайде изображена корзина, в которой и рядом с которой были свежие грибы. Или берег моря (озера) с людьми, одетыми в купальные костюмы. Вопрос: «В какое время года мог быть сделан этот снимок?» Правильный ответ для наших широт: «Летом». Другой подход к той же проблеме – описываются так называемые чувственно-конкретные признаки предмета: «овальный», «кислый», «желтый». Вопрос: «Что это?» Ответ: «Лимон» (или «слива») и т. п. (Бундзен, Давид, 1979).
Естественно, здесь приведены не все использованные тесты, а их основные типовые варианты.
Импульсная активность нейронных популяций записывалась с тех зон мозга, куда по диагностическим и лечебным соображениям вводили долгосрочные электроды. Однако индивидуальные вариации строения мозга определяли необходимость и целесообразность введения несколько избыточного количества электродов. Так, при необходимости введения электродов в мозговое образование, по протяженности (вертикали, горизонтали или другой плоскости) не превышающее в типовом варианте одного сантиметра, электродный пучок обеспечивал возможность регистрации физиологической активности на протяжении от полутора до двух сантиметров дискретно, через каждые 2–3 мм межэлектродного расстояния в пучке. С диагностических и лечебных позиций такая избыточность обеспечивала при всех индивидуальных вариациях достаточно надежное взаимодействие с заданной структурой. В научном плане, хотя введение электродов им и не определялось, при этом практически всегда оказывалось возможным регистрировать физиологическую активность со структур мозга и их зон, пограничных с целевой зоной. Таким образом оказалось исследованным очень большое количество точек мозга – порядка нескольких тысяч – у более чем ста больных паркинсонизмом, с фантомно-болевым синдромом, гепатоцеребральной дистрофией (болезнью Вильсона–Коновалова), эпилепсией, а позднее – с опухолями и травмой черепа и головного мозга. Точки располагались в коре, верхних отделах ствола, ядрах стриопаллидарной системы, таламуса и в образованиях медиобазальных отделов височной доли. Как указывалось выше, электроды вводились по клиническим показаниям в зоны мозга, предположительно важные для диагностики и лечения. Для точного типового введения по стереотаксическим атласам использовались возможности ЭВМ. При паркинсонизме и гепатоцеребральной дегенерации электроды вводили преимущественно в таламические ядра, образования стриопаллидарной системы, верхние отделы ствола и премоторную кору. При этом своего рода обязательными мишенями были вентролатеральный комплекс и срединный центр зрительного бугра, медиальный членик бледного шара, хвостатое ядро, верхние отделы ствола и премоторная кора. При фантомноболевом синдроме наряду с этими зонами электроды вводили целенаправленно и в более дорсально расположенные ядра зрительного бугра, и в подушку. При эпилепсии практически во всех случаях их вводили в медиабазальные отделы височных долей (крючок извилины гиппокампа, различные отделы гиппокампа, миндалину), в вентролатеральный комплекс и срединный центр зрительного бугра,.передние и медиальные ядра зрительного бугра, хвостатое ядро, перегородку, верхние отделы ствола, поясную извилину, височную кору и реже – в другие области мозга в зависимости от особенностей заболевания у конкретного больного. Принципиально новыми в последние годы являются исследования различных зон конвекситальной коры. Данные о локализации электрода предварительно получали на основе расчета с помощью ЭВМ по стереотаксическому атласу с внесением индивидуальной анатомической коррекции (Аничков и др., 1976; Аннарауд, Усов, 1976; Аничков, 1977; Аничков, Полонский, 1977; Полонский, 1977). Реальное расположение электродов в мозгу оказывается, как правило, совпадающим с заданным или очень к нему близким.
Эти сведения, однако, отражали индивидуальные вариации лишь в общем виде. Для использования вживленных электродов с лечебной целью необходимо иметь более надежные критерии прижизненного определения их точной локализации. Этим требованиям в наибольшей мере отвечают данные стереотаксической неврологии, базирующейся на тонком исследовании эффектов электрической стимуляции через вживленные электроды в сочетании с результатами других приемов комплексного метода. Однако для правильного использования данных стереотаксической неврологии необходимо учитывать физиологические (патофизиологические) условия внутримозгового взаимодействия, зависящие от характера заболевания. Так, наиболее точно судить о расположении электрода по данным стереотаксической неврологии, то есть по характеристике эффектов, развивающихся при точечном электрическом воздействии через вживленные электроды, можно при паркинсонизме в связи с несколько ухудшенными условиями внутримозгового взаимодействия (Петушков, 1970, 1977, и др.). Наоборот, при эпилепсии эти данные нуждаются в особо тщательном анализе для дифференцирования местных и дистантных эффектов, развивающихся в связи с возможностью общего или частного облегчения внутримозгового взаимодействия (Бехтерева и др., 1978). Сказанное, естественно, не должно восприниматься как дискредитация возможностей стереотаксической неврологии в целом, ибо с учетом поправок на особенности заболевания это направление в физиологии мозга человека является новой ступенью познания его структурнофункциональной организации.
При исследовании нейрофизиологических коррелятов фонем, слогов, слов сотрудники лаборатории вполне отдавали себе отчет в том, что изучаемые нейронные популяции находились вне классических слуховых образований мозга, обычно являющихся объектом анализа при предъявлении акустических сигналов. В то же самое время опорные физиологические данные, свидетельствовавшие об изменении физиологических показателей мозга в этих структурах при вербальных психологических тестах, подкрепленные анатомическими данными о наличии связей этих структур со слуховыми образованиями мозга, а также данными С. Б. Дзугаевой (1975) о прямых непереключающихся путях от рецепторных приборов к различным внутримозговым зонам, определяли правомерность использования фонограмм не только как ориентира предъявления и воспроизведения сигнала. Все это позволило предположить, что акустические характеристики используемых в тестах словесных сигналов и их элементов могут отражаться в перестройках импульсной активности нейронных популяций, хотя, естественно, трудно было ожидать прямого и достаточно полноценного отражения именно этих признаков.
Перестройка импульсной активности при предъявлении гласных фонем, произносимых исследователем, у разных больных проявлялась в области бледного шара, срединного центра, вентролатерального ядра (Перепелкин, 1974, 1977). С несколько другими целями (для изучения мозговых механизмов простейших видов принятия решения) сотрудница лаборатории Н. Г. Шкурина (1978) применяла в тестах искусственные фонемы [А], [Э], [И], синтезированные в Швеции в лаборатории Фанта и предоставленные в наше распоряжение Л. А. Чистович. Основные результаты сводились к тому, что в различных областях срединного центра, вентролатерального ядра, бедного шара и других зон обнаруживались более или менее устойчивые во времени пространственно-временны?
е перестройки импульсной активности, позволяющие дифференцировать фонемы и слоги. В дальнейшем исследование было продолжено при использовании показателя динамики формирования и воспроизводимости групповых последовательностей разрядов с определенными интервалами. Специальное внимание уделено увеличению связности импульсного потока и конкретно соотношению триад в фоне и при реализации тестов, различию перестроек импульсной активности в разные фазы теста (восприятие фонемы, произнесение про себя, произнесение вслух) и в зависимости от использования в тесте отдельных фонем, слогов, значимых и незначимых триграмм и слов с учетом объективной и субъективной вероятности их появления в речи. Исследована также зависимость перестроек импульсной активности от акустических характеристик сигнала. Исследования показали, прежде всего, наличие хотя и не простой, но достаточно отчетливой связи между составляющими импульсного паттерна и акустическими характеристиками входного сигнала. П. Д. Перепелкин при предъявлении русских фонем и слогов в стриопаллидарной системе наблюдал последовательности разрядов с определенными интервалами, селектировавшиеся перед произнесением больным соответствующих гласных (исследования проводились в реальном времени). Н. Г. Шкурина (1978) наблюдала в премоторной зоне коры и в области бледного шара появление триад, усредненные значения которых различались для синтетических фонем [А], [Э], [И] (табл. 2).
Таблица 2
Усредненные длительности межимпульсных интервалов в триадах, общих для каждого из эталонных стимулов, и периода идентификации стимула по нейронным популяциям премоторной области коры (I, II) и бледного шара (III )
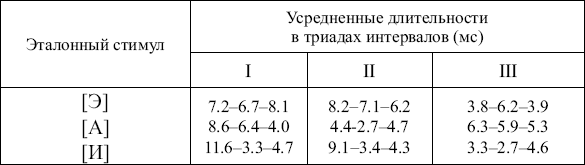
В исследованиях с предъявлением синтетических фонем (Шкурина, 1978) изучались совокупности триад, формирующихся и воспроизводящихся во время теста в мультиклеточной активности нейронных популяций в ответ на предъявление синтетических гласных. Для этой работы характерно было изучение не избранных, проверенных на основе эталонного поиска триад, а всех, обнаруженных в мультиклеточной активности во время этого теста. Логично было бы предположить, что наряду с характерными для данной фонемы при такой постановке исследования в обследованных зонах могли обнаруживаться не только «необходимые», но и «избыточные» мозговые перестройки. Однако и на основе сравнения усредненных триад дифференциация фонем по мозговым перестройкам оказалась возможной. Предъявление смешанных синтетических фонем, которые могли бы распознаваться и как [А], и как [Э], показало, что с 60%-ной вероятностью на основе возникающих в мозгу триад возможно предсказать, как именно будет опознана фонема. Исследования с изучением триад, групповых последовательностей четырех разрядов соответственно с тремя интервалами определенной продолжительности (допуск +10 %) обнаружили, что в групповых последовательностях разрядов в определенный период функционально значимым оказывался именно параметр длительности интервала между разрядами, а не образующие эти интервалы разряды нейронов.
В ряде нейронных популяций, в которых обнаруживалась реакция различных физиологических показателей мозга на вербальные психологические тесты, в ответ на произнесение известных и неизвестных, частых и редко встречающихся в речи слов и словоподобных образований из фонем родного языка отмечалась пространственно-временна?я перестройка импульсной активности нейронной популяции – формирование паттерна (Бехтерева и др., 1977а). Как указывалось выше, удобно для использования в качестве эталона для введения в ЭВМ и достаточно полно для последующего исследования этот паттерн представлять в виде последовательностей интервалов между разрядами группы нейронов – в дистрибутивной форме.
Исследования динамики этого паттерна обнаружили ее зависимость от фазы теста, от частоты встречаемости слова в речи; динамика была различной при предъявлении знакомых и незнакомых слов и словоподобных сигналов. Характер паттерна мог быть различным в зависимости от задачи, в процессе которой воспринималось, удерживалось в памяти и воспроизводилось слово, или от ситуации контекста, в связи с которым паттерн слова извлекался из памяти при более сложных психологических тестах. При предъявлении в тестах известных слов паттерн первоначально обнаруживался в полной, развернутой форме, по времени сопоставимой с услышанным словом. Непосредственно после окончания звучания слова и исчезновения из импульсной активности полного паттерна он вновь появлялся, но уже в компрессированной форме. Это явление еще полностью не изучено. Судя по первоначальным данным, происходит трансформация первого паттерна с сохранением его основных опорных значимых элементов. Затем паттерн исчезал из импульсной активности и появлялся вновь непосредственно перед произнесением слова. Попытка сопоставления первоначально возникшего и развивающегося перед ответом паттерна с акустическими характеристиками входного и выходного сигналов – слов, произносимых врачом и больным, показала, что в первом паттерне находили отражение характеристики ча стотно-амплитудной огибающей спектра слов, произносимых врачом, во втором – слов, произносимых больными. Эти данные более подробно представлены в одном из наших предыдущих обобщений (Бехтерева и др., 1977).
Использование паттернов в качестве эталонов при машинном анализе подтвердило значение перестроек, развивающихся в мозгу перед произнесением слова: эталон был более надежным, если в него включалась не только та перестройка, которая наблюдалась в момент отлетной реакции, но и та, которая развивалась примерно за 40±25 мс до нее (Бундзен и др., 1975б). Подтверждением физиологического значения этого периода служили и данные, полученные при изучении мышечных механизмов словообразования и импульсной активности отдельных нейронов перед вербальным ответом (см. выше). Наличие перестроек, предшествующих ответу, с учетом приведенных подтверждений дает известные основания предполагать существование корреляции мозговых паттернов не только с акустическими, но и с моторными характеристиками слова. Полностью признавая гениальность сеченовской мысли о значении отражения моторных факторов в мозговой деятельности, в том числе психической, мы полагаем, что в мозговом паттерне наряду с акустическими характеристиками входного и выходного сигнала находят отражение и моторные характеристики слова. Паттерн формируется на основе ряда факторов, в числе которых сенсорным и моторным должна быть отведена ведущая роль, хотя ими не исчерпывается перечень определяющих если не формирование, то динамику паттерна причин.
Предъявление больному неизвестных иностранных слов, где незнакомыми были и бо?льшая часть составляющих их элементов, и значение самого слова, также приводило к формированию паттерна, однако динамика его в последующем была отличной от возникающей при отражении знакомых слов родного языка (Гоголицын, 1973а, 1974). Возникший первоначально паттерн далее, хотя и в дискретной форме, обнаруживался до воспроизведения больным заданного слова. Это явление было обозначено по контрасту с тем, что наблюдалось при предъявлении известных слов, как ультрастабильность паттерна (хотя приставка «ультра» относилась скорее к эмоциональной реакции исследователей на этот факт, чем к реальному положению дел). На этом фоне не удавалось выделить новый управляющий паттерн: управлению ответом служил, по-видимому, первоначально образовавшийся и удерживающийся далее паттерн. Теперь, уже на новом уровне изучения мозговых паттернов, вопрос нуждается в дополнительном исследовании деталей, однако первоначально установленная зависимость паттерна от базиса долгосрочной памяти, сформировавшейся в результате индивидуального опыта, подтверждается. Паттерн, возникший при восприятии слова, которое может быть опознано, которому есть аналог в долгосрочной памяти, исчезает из импульсной активности вскоре после появления и возникает вновь в трансформированном виде к моменту ответа. Паттерн незнакомого слова сохраняется до ответа. При этом стабильный паттерн как бы выполняет по крайней мере двойную службу: он не только становится мозговой основой формирования ответной реакции, но и формирует соответствующий базис долгосрочной памяти. Следует подчеркнуть, что эти особенности динамики паттерна обнаруживались только в тестах на удержание задания в памяти с последующим повторением его по просьбе врача, то есть при соответствующей мотивации.
При изучении этой кратковременной стабильности мозговых паттернов в зависимости от степени известности слова должны варьировать не только словесные сигналы, но и условия исследования, детали задания. Подтверждением зависимости динамики паттерна от базиса долгосрочной памяти служит ее перестройка после того, как иностранное слово в процессе повторения и обучения становится знакомым больному. Предъявление в тесте на краткосрочную память того же самого, ранее неизвестного, а теперь знакомого слова приводит к возникновению паттерна, его быстрому исчезновению и новому появлению в форме, аналогичной для ранее обнаруженного управляющего паттерна. При этом характерно, что уже в момент предъявления иностранного слова в паттерне, соответствующем фазе восприятия сигнала, обнаруживаются признаки, содержащиеся в характеристиках паттерна, возникающего на то же слово на родном языке. Так, например, в тесте использовалось немецкое слово «bogen». «Bogen», «bogen» – «повторите» – произносится что-то более или менее близкое к этому слову. Больному поясняли, что слову «bogen» наиболее соответствует русское слово «арка». Соотношение bogen–«арка» легко заучивалось. Далее в фазе, соответствующей восприятию уже теперь известного слова «bogen», не содержащего, как известно, фонемы [А], дважды повторяющейся в русском слове «арка», даже применявшимися ранее относительно грубыми приемами анализа мультиклеточной активности (в частности, способом факторного анализа фрагментов текущей частоты) улавливались моменты появления соответствия коррелятов фонемы [А], свойственной русскому переводу. Больной не знал немецкого языка. Он выучил только одно слово. Естественно, что уже при начале опознания этого иностранного слова он переводил его на родной язык, признаки чего и обнаруживались в перестройках мультиклеточной активности (Гоголицын, 1976а). Вполне понятно, что интересным и необходимым для дальнейшего изучения нейрофизиологических коррелятов психической деятельности этапом было исследование отражения в перестройках импульсной активности смысловых характеристик слов. В психологических исследованиях такие задачи решались без учета внутримозговой нейродинамики (Лурия, Виноградова, 1971). В более поздних, чем наши, нейрофизиологических работах Браун и соавторы (Brown et al., 1976) строят фразу, используемую в качестве теста таким образом, что слово, нейрофизиологические корреляты которого исследуются, стоит всегда на определенном месте внутри предложения, в частности, является последним. В зависимости от предшествующего ему текста меняется смысл этого слова. При этом улавливается разница в динамике вызванных потенциалов с поверхности кожи черепа. При таком подходе в динамике вызванного ответа определяются корреляты не смысла слова, а дифференциальных признаков одинаково звучащих, но разных по смыслу слов. По-видимому, сегодня наиболее целесообразны различные вариации именно таких исследований, основанных на поисках нейрофизиологического выражения смысловой общности или смысловой дифференциации, хотя не исключены и другие подходы. Это направление разрабатывается также в исследованиях Джона (John, 1976, 1977), хотя вряд ли опознание знака I, предъявленного на слайде как цифры или буквы в зависимости от контекста (1, 2 или I, J), по характеру вызванных потенциалов давая статистически результаты, удовлетворяющие требованиям нейрометрики, существенно более близко труднее накапливаемым данным о нейрофизиологических коррелятах собственно семантики слов. Различия в вызванных потенциалах в зависимости от значения сигнала исследуют Гукер (Gucker, 1973), Ю. Г. Кратин (1976), а также А. А. Пирогов, И. Месарош (1976), Т. П. Зеленкова, А. М. Иваницкий (1979) и многие другие.
Продолжаются попытки использовать для изучения психической деятельности ЭЭГ (Русаков, Бодунов, 1977; Хризман, Зайцева, 1977, и др.). Изменение мозгового кровотока при психической деятельности с помощью приема многоканальной записи изучает в течение многих лет Ингвар (Ingvar, 1976).
В работах Джона (John, 1977) также показано, что в нейронных ансамблях могут развиваться воспроизводимые паттерны, различные на разные раздражители, и, таким образом, приведена возможность наблюдать отражение значения сигнала и в импульсной активности нейронов. По данным Хелгрина с соавторами (Halgren et al., 1978; Ravagnati et al., 1979), 33 нейрона в извилине гиппокампа строго избирательно реагировали на определенное содержание разговора с больным и молчали, если разговор касался других тем. В работах нашей лаборатории использовалось несколько вариантов психологического подхода к вопросу и соответственно несколько вариантов опорного анализа нейрофизиологических данных.
Оказалось, что обнаруживать нейрофизиологические признаки смысловой общности слов и дифференциальные признаки разных смысловых полей возможно по данным импульсной активности нейронных популяций при предъявлении слов общего для них смыслового поля, то есть понятий, имеющих такую смысловую взаимосвязь, которая позволяет обобщать их каким-то одним словом. Типовые варианты тестов приведены выше. Уже первоначальные опорные исследования, проведенные способом факторного анализа, показали, что в нейрофизиологическом паттерне, развивающемся (Гоголицын, 1976б) при последовательном предъявлении таких серий слов в медиальном членике бледного шара, в хвостатом ядре и центральном покрышечном пути, обнаруживаются характеристики, общие с паттерном, регистрируемым при восприятии и произнесении слова-обобщения. При этом в некоторых случаях анализ нейрофизиологического паттерна обнаруживал иерархическую организацию, иерархические признаки общности. Так, например, если задание состояло из длинного ряда названий деревьев, обнаруживалось сходство паттернов, возникающих в ответ на названия отдельно хвойных и отдельно лиственных деревьев, и более общее сходство паттернов с паттерном на слово «деревья». С использованием отдельных элементов паттерна, групповых последовательностей разрядов с определенными интервалами эти данные не только подтверждались, но и оказался возможным их автоматизированный контроль способом эталонного поиска.
В. Н. Малышев (1977) на основе анализа импульсной активности, зарегистрированной в период теста на обобщение, построил двухмерные матрицы, где по одной оси были нанесены элементарные составляющие нейрофизиологических паттернов, а по другой – предъявляемые слова, не имеющие скольконибудь существенной акустической общности, но объединяемые по смыслу. В связи со сложностью анализа в матрице была приведена часть характерных для этих слов нейрофизиологических элементарных составляющих паттерна, зарегистрированного в области различных ядер зрительного бугра, стриопаллидарной системы и верхних отделов ствола. Показано, что имелись элементарные составляющие паттерна – групповые последовательности разрядов с определенными интервалами, которые обнаруживались у всех или почти у всех слов данного смыслового поля. При использовании такой элементарной составляющей в ЭВМ в качестве эталона и последовательного анализа записей фоновой импульсной активности нейронных популяций, активности, зарегистрированной при выполнении больным данного теста и аналогичного теста, состоящего из слов другого смыслового поля, машина «выбирала» слова своего смыслового поля. Эти результаты целесообразно сопоставить с экспериментальными исследованиями (Толкунов, 1978), в которых наблюдалась перестройка интервальной структуры нейронной активности в зависимости от информационного содержания сигнала.
Не было сомнений в том, что важнейший вопрос об отражении смысла слов в характеристиках паттерна нуждался в дальнейшем изучении. Развитие исследований пошло по пути накопления данных и их анализа, адекватно стандартизированного с помощью необходимого математико-технического аппарата, а также разработки новых модификаций психологических подходов к вопросу.
Иногда еще, хотя теперь и реже, встают вопросы о том, можно ли в структурах стриопаллидарного комплекса, зрительного бугра, миндалины, гиппокампа, верхних отделов ствола и коры обнаружить зоны, отражающие характеристики слова и психической деятельности. Выше приведены экспериментальные физиологические и морфологические данные, а также результаты, полученные другими учеными в исследованиях, проводимых у человека, прямо или косвенно свидетельствующие о целесообразности и важности изучения этих вопросов, предпосылкой постановки которых был длительный период изучения нами структурно-функциональной организации психической деятельности. Интересно также выяснить, как в количественном выражении распределяется в пределах структур, куда по клиническим показаниям вживлялись электроды, число зон мозга, где реакция импульсной активности на вербальные тесты обнаруживается в неспецифической и специфической (паттерн и его элемент) формах. Несмотря на всю важность привязывания динамики к структуре, много лет назад задачей первоочередной важности было прежде всего выявление зон, где обнаруживались соотношения с акустическими и семантическими свойствами словесных сигналов. В публиковавшихся статьях и по ходу обобщения вопроса в монографии Н. П. Бехтеревой с соавторами (1977а) указывалось, где обнаружены корреляты со смысловыми и/или акустическими характеристиками слов, причем не всегда отмечалось, что из числа обследованных п зон х структуры отражение смысловых (акустических) характеристик слова и т. п. обнаружено т раз. Сегодняшний период исследования позволяет представить, пусть предварительно, и такого рода данные, хотя в этом случае зоны, попавшие в число п–т, должны еще далее изучаться. Для того чтобы четко сказать в отношении их «нет», нужны годы исследований, в то время как т—«да» – накапливаются по ходу работы ежедневно.
Что дают эти данные для нейрофизиологии психической деятельности человека? Число обследованных структур мозга не мало, но не исчерпывающе. В нем не представлены еще с необходимой полнотой многие образования коры больших полушарий. Мало исследованы важнейшие для нормального течения психических процессов медиобазальные отделы височных долей. На основании имеющихся данных нельзя судить во всей полноте о системе обеспечения психической деятельности. И все же уже и эти данные свидетельствуют о том, что глубокие структуры мозга участвуют в качестве звеньев в системе обеспечения психической деятельности, причем звеньев не только энергетических, но, повидимому, и информационных (!), а также о том, что далеко не во всех исследованных структурах обнаруживаются тонкие изменения нейрофизиологических процессов при психической деятельности. Располагая этими материалами, мы уже сегодня можем возражать против различных вариантов все еще распространенного мнения о том, что мозг принимает участие в обеспечении психической деятельности в целом (Pribram, 1975; John, 1977, и др.). В пользу наших представлений говорят и данные ряда других исследователей (Лурия, 1977; Ojemann, 1979, и др.).
В сравнительно ранних, упоминавшихся выше работах говорилось о том, что, когда больному называли неизвестное ему иностранное слово, а затем вновь повторяли его после перевода этого слова на русский язык, наблюдалась не только перестройка динамики нейрофизиологического паттерна. В процессе этих исследований был использован следующий вариант теста: больному называлось немецкое слово, а ответ предлагалось произносить по-русски. Больной выполнял задание. Но в той же нейронной популяции срединного центра зрительного бугра, где паттерн ранее обнаруживался, он не появился вновь ни перед ответом на русском языке, ни во время ответа! (Гоголицын, 1974). Этот факт может лечь в основу исследований различий структурнофункциональной организации психических процессов, реализуемых на основе родного и иностранных языков. Было бы ошибкой делать далеко идущие выводы из неполных материалов. Однако начинают появляться исследования, подтверждающие эти данные о наличии у мозговых систем, обеспечивающих возможности мышления на разных языках, и общих, и различных звеньев (Ojemann, Whitaker, 1978a, 1978b).
Эти данные были получены авторами во время психологического теста, проведенного по ходу электрической стимуляции мозга. Предъявлялись слайды с изображением предмета, который люди, владеющие двумя языками, должны были назвать на обоих этих языках. При стимуляции центральных речевых зон нарушалась возможность называния слов на обоих языках. На периферии, в лобной и теменной коре, были точки, имеющие отношение только к одному языку! Структурно-функциональная организация мозговых систем, обеспечивающих мышление на родных языках, была различной у двух обследованных больных. Может быть, именно на этом пути лежит ключ к пониманию интереснейших клинических фактов – случаев внезапного забывания одного из языков, в том числе родного, при некоторых поражениях мозга. Пока не накоплен достаточный опыт, можно сказать, что в некоторых зонах мозга характер протекающих нейрофизиологических процессов определяется только акустическими свойствами слов. В специально ориентированных исследованиях Ю. Л. Гоголицына, проведенных в условиях «ожесточенной» достоверности способом постстимульных гистограмм, показано, что обнаруживаются зоны мозга, в динамике импульсной активности которых не получают отражения различия слов с одинаковым звучанием. Такие зоны выявлены в том же хвостатом ядре, вентролатеральном ядре таламуса, а возможно, будут обнаружены и в других структурах. Мы еще не можем утверждать, что для зон, где обнаруживаются нейрофизиологические признаки смысловой дифференциации, акустические характеристики не имеют значения. На сегодня рабочая гипотеза может быть сформулирована так: в ряде зон мозга, не являющихся классическими слуховыми образованиями, при восприятии, удержании в памяти и воспроизведении слов формируются паттерны, в которых отражаются акустические и смысловые характеристики слова, по-видимому, его моторные характеристики, а сам паттерн и его динамика подчинены базису долгосрочной памяти, сформировавшемуся в результате индивидуального опыта.
Уже в самых первых публикациях, посвященных расшифровке процессов, протекающих в подкорковых и некоторых корковых звеньях системы обеспечения психической деятельности при ее реализации, нами было использовано понятие «коды». Надо сказать, что в смысле введения этого термина в нейрофизиологию мы не были пионерами. Термины «коды» и «кодирование» используются многими исследователями, причем нередко в тех случаях, когда кодирование сигнала в нервной системе происходит или должно происходить, но именно это явление не исследовано. Большой обзор данных по кодированию сигналов в периферических и центральных отделах нервной системы сделан Сомьеном (Somjen, 1975). Анализ состояния проблемы по нашему предложению проведен П. В. Бундзеном, Ю. Д. Кропотовым и В. В. Трубачевым (1978). Известно, что если в периферических отделах анализаторных систем обнаружены более или менее сопоставимые с сигналами внешнего мира показатели, то в головном мозгу высокоорганизованных животных и особенно человека трансформации сигнала той степени тонкости, которая могла бы претендовать на это определение, до последнего времени прослежено не было.
В книге В. Е. Анисимова (1978) приведен ряд определений понятия «коды» и «кодирование». Не со всеми определениями можно согласиться, но данный перечень свидетельствует о множестве смысловых оттенков, которые вкладываются в эти понятия. При этом утверждается, что нет и принципиально не может быть единого преобразования (кода) сигнала на разных уровнях в мозгу – органе, в котором равно неправомерно недооценивать роль структуры в реализации функции и роль нейродинамики в структуре при протекании того же процесса. Основные из этих определений приводятся ниже.
1. В процессе хранения, передачи и обработки информации требуется преобразование одного сигнала в другой, наиболее пригодный для выбора канала изменения формы сигнала; установление определенного соответствия между сообщением и той системой символов, при помощи которых эти сообщения передаются. Все эти операции называются кодированием.
2. Под кодированием понимается определенная операция шифрования информации, которая превращает сообщение в сигнал. При этом содержание информации не изменяется, оно лишь принимает иную форму. Кроме того, чтобы закодированные сигналы являлись переносчиками информации, всегда избирается такая система кодирования, которая позволяет их обращение, то есть декодирование. Декодирование сигналов означает восстановление начального сообщения.
3. Под кодом понимают в частном случае определенную систему символов.
4. Выбор наиболее экономичного кода обеспечивает быструю передачу сообщения, информации, однако при этом вероятность искажения сигнала увеличивается. Чтобы уменьшить эту вероятность, передают избыточную информацию.
5. Кодированием называется процесс преобразования одного сигнала в другой, наиболее пригодный для передачи информации по конкретному каналу связи, – процесс установления определенного соответствия между сообщением и той системой символов, при помощи которых эти сообщения передаются.
6. В более широком смысле под кодированием понимается любое сопоставление, в том числе одних знаков или сигналов с другими.
Этими формулировками понятие «кодирование» не исчерпывается. Тех, кто дополнительно интересуется именно терминологической стороной проблемы, отсылаем к книге Сомьена (Somjen, 1975).
К какому классу процессов целесообразнее всего отнести паттерны, описанные в наших статьях, монографии и в данной книге? Они безусловно подходят под всеобъемлющее понятие «коррелят», однако именно в данном случае это понятие в связи с тем, что обычно вкладывается в указанный термин, не отражает уровня изучения вопроса. Паттерн, выделенный из импульсной активности, обладает той степенью характерности, которая позволяет использовать его в качестве эталона при эталонном машинном поиске аналогов этого паттерна в импульсной активности. Нейрофизиологические корреляты, как правило, менее специфичны и, отражая в нервной системе какое-то конкретное явление, за очень редким исключением, по существу, отражают класс явлений. Так, например, местные медленные волны на ЭЭГ указывают на изменения состояния нервной ткани, развивающиеся при опухолях мозга. Но если заложить в качестве эталона в ЭВМ отрезок записи медленноволновой активности, то окажется, что машина выловит из ЭЭГ сообразно с эталоном медленные волны и при опухолях, и при очаговых сосудистых поражениях, и при травме, и при воспалительном процессе, и во время сна здорового человека. Более специфичным окажется в качестве эталона наблюдающийся при эпилепсии комплекс острой и медленной волн (спайк–волна). В этом случае машина быстро обнаружит его аналоги, и без того хорошо видные на ЭЭГ невооруженным глазом.
Однако в случае тех паттернов, о которых идет речь, заложив один из них в ЭВМ, на отрезке записи от десятков секунд до десятков минут можно опознать момент появления именно его аналога в импульсной активности, например, заложив эталон, соотносимый с другим словом, импульсную перестройку, сопоставимую с этим другим словом, и т. д. Паттерны оказываются приемлемыми для использования в качестве эталонов при машинном поиске, они различны для разных групп слов, и, таким образом, если рассматривать эти паттерны как корреляты слов, приходится признать, что это корреляты более высокого, чем обычно, порядка. Но, как показано выше, паттерн, выделенный из импульсной активности, может быть расщеплен на составляющие части, по крайней мере, часть из которых обладает достаточной характерностью для использования в качестве эталона при машинном поиске аналогичных паттернов в импульсной активности нейронных популяций. Таким образом, паттерн все же достаточно характерен, как и некоторые его элементы. При использовании такого элемента в качестве эталона в импульсной активности обнаруживаются те паттерны, которые имеют в своем составе данный элемент и соотносятся с отражением в мозгу определенных слов. Используя этот эталон при анализе фоновых записей, естественно, выявляли единичные аналоги его, но анализ записей импульсной активности, осуществленных при восприятии, удержании в памяти, воспроизведении и обобщении других слов, с помощью данного эталона был малоэффективен.
Рассматривая данные анализа мультиклеточной активности, следует разобраться, почему именно триада рассматривается как единица паттерна. Действительно, в импульсной активности выделяются и другие регулярные элементы – повторяющиеся последовательности с меньшим или большим количеством определенных интервалов. Так, например, в импульсной активности парных интервалов наблюдается значительно больше, чем триад.
При одновременной обработке достаточно большого количества нейронов из активной популяции оказывается, что и в фоне (редко), и в момент теста обнаруживаются воспроизводимые групповые последовательности разрядов, но уже не с тремя, а с бо?льшим числом определенных интервалов. И все же есть ли достаточно убедительные основания для того, чтобы считать именно триаду основной или единственной элементарной единицей паттерна? Единственной – нет.
Паттерн может содержать и связанные пары интервалов, и большие регулярные группы разрядов. Элементарной единицей паттерна может быть и последовательность из четырех и более интервалов. Что, однако, важно подчеркнуть при рассмотрении данного вопроса – именно триада может быть минимально достаточной характерной единицей паттерна. Именно триады являются и более устойчивыми элементарными единицами, хотя это не исключает значения элементов с большим количеством последовательных разрядов.
В паттерне, соотносимом со словом или его элементом, как указывалось в ряде проведенных нами исследований, может быть обнаружено множество триад. Проверить степень специфичности каждой из них, время устойчивости первоначально возникшего множества элементов паттерна нелегко, но возможно с помощью уже имеющихся методов. В работах Н. Г. Шкуриной, исследовавшей нейрофизиологическое отражение синтетических фонем, показано, что и этот полный комплекс – паттерн – может быть использован для дифференцировки отражения разных фонем. Следует помнить, что отдельные триады при эталонном поиске, наоборот, могли быть использованы для выделения группы слов общего смыслового поля. Отсюда, если целесообразна проверка значимости и характерности отдельных элементов, то целесообразна и своего рода обратная операция – исследование динамики характерности паттерна при последовательном изменении количества триад, вводимых в паттерн и выводимых из него. Именно на этих путях лежит решение вопроса о том, какой набор элементов в одной нейронной популяции является необходимым и достаточным отражением одного слова. Однако, зная хотя бы некоторые из общих механизмов мозга, следует учитывать, что избыточность не означает ненужности.
Далее, как указывалось выше, характерные перестройки импульсной активности – паттерны – наблюдаются в исследуемых условиях не в одной, а в нескольких популяциях, в нескольких зонах мозга, а если учесть условия исследования, повидимому, правомерно сказать – во многих зонах. Как соотносятся паттерны разных зон мозга?
Организация импульсной активности в одних и тех же условиях варьирует в разных зонах мозга и в так называемой фоновой записи – до предъявления теста, и при восприятии слов, словоподобных сигналов, слогов и фонем. Выделение из общего паттерна его элементарных составляющих показывает, что большая часть их оказывается различной в разных зонах мозга. Однако имеются и сходные по последовательным межимпульсным интервалам группы. Рабочей гипотезой (Бехтерева, 1977), объясняющей на сегодня эти факты, является представление о распределенности в мозгу полного паттерна – отражения – и о значении дублирующих элементов как системообразующих, организующих систему факторов. Эта гипотеза может проверяться прежде всего путем исследования функционального значения составляющих паттерна методом эталонного поиска. Однако и уже имеющиеся материалы по динамике паттерна в процессе более сложной психической деятельности заставляют предположить, что дело, особенно в отношении синхронных элементов в разных зонах мозга, обстоит если не иначе (для такого мнения нет оснований), то сложнее.
Таким образом, в нашем случае использование терминов «код» и «кодирование» основано прежде всего на характерности выделяемых короткоживущих паттернов или по крайней мере некоторых их элементов, позволяющей на определенных отрезках времени не только наблюдать появление этой перестройки в мозгу при одном и том же внешнем сигнале (слове и т. д.), но и определять в импульсной активности моменты возникновения соответствующих отражений слов, словоподобных сигналов и их элементов, используя предварительно выделенный их аналог в виде эталона при основанном на сравнении с этим эталоном машинном поиске. Фактами, на которых основывается представление о кодовом значении регистрируемых паттернов, является и функциональная значимость их элементов, и возможность встраивания элементов в паттерн в специальных условиях исследования. По-видимому, и сама динамика паттерна по ходу простейших тестов на восприятие и воспроизведение слов свидетельствует также о кодовом значении этих паттернов: паттерн возникает в момент восприятия слова, далее претерпевает различные трансформации в зависимости от базиса долгосрочной памяти и вновь появляется перед произнесением и во время произнесения слов. Кодовое значение паттернов подчеркивается наличием в паттернах разных зон мозга сходных элементов. И в то же самое время использование в данном случае «кодовой» терминологии требует во избежание недоразумений пояснений и постановки ряда вопросов, решение которых важно для дальнейшего изучения проблемы мозгового кода психической деятельности.
Так, и это, пожалуй, самое основное – исследование активности многих нейронных популяций подчеркнуло, что регистрируемые паттерны – это местные явления в мозгу. Во всяком случае паттерн в таком виде не передается с соответствующих анализаторных, в данном случае слуховых, образований мозга, а возникает местно под влиянием факторов, рассмотренных выше. В таком полном виде, по-видимому, паттерн не передается и дальше. Это положение является не только результатом логического сопоставления массивности и сложности данного элемента с передаточными возможностями каналов связи и временны?ми параметрами очень быстрых процессов – психических явлений, но и тех же исследований паттернов в большом числе разных нейронных популяций мозга. Обнаруженные и исследованные нами паттерны – местные, привязанные к структуре тончайшие нейрофизиологические корреляты психической деятельности или, точнее, нейрофизиологические выражения кодирования и декодирования, детерминированные в значительной мере памятью и обучением. Это – та нейродинамика (или ее часть), возникновение которой находится в прямой зависимости от участия структуры в обеспечении психической деятельности. Естественно, очень важно знать, как, на основе каких процессов осуществляется взаимодействие между структурами в высших отделах нервной системы? Но это уже другой вопрос, хотя и связанный с проблемой местного кода. Он будет рассмотрен нами далее.
Пока еще трудно с уверенностью сказать, какой именно уровень интеграции обеспечивается деятельностью исследуемых структур стриопаллидарной системы, зрительного бугра, верхних отделов ретикулярной формации (весьма, кстати, дифференцированной – П. Г. Костюк, 1973), медиобазальных отделов височной доли, премоторной и некоторых других отделов коры. Как будто трудно отрицать на основе все увеличивающегося числа экспериментальных функционально-анатомических данных и исследований, проводимых у человека в разных лабораториях мира, что изучаемые подкорковые зоны мозга значительно более специфичны, чем это предполагалось ранее, и если не являются единственными областями высшей интеграции при осуществлении психической деятельности, то, безусловно, в ней очень активно участвуют (Костандов, 1978, и др.). В рецепторных и анализаторных образованиях идет первичное кодирование сигнала, перекодирование, адаптированное к передаче, перекодирование для первичного анализа. Этот вопрос подробно рассматривается в работах Уордена и Марша (Worden, Marsh, 1968), Я. А. Альтмана (1972), Кейдела (Keidel, 1974), Моллера (Mоller, 1974, 1978а, 1978б), Михольсона и Винсента (Michelson, Vincent, 1975), Десмедта (Desmedt, 1977), Мушегиана с соавторами (Moushegian et al., 1978), Л. А. Чистович (1978) и ее сотрудников и многих других. В высших по отношению к анализаторным структурах, в которые стекается информация от разных анализаторов по прямым и непрямым путям, происходит или, точнее, может происходить расшифровка свойств более сложных сигналов, в данный момент или онтогенетически воспринимавшихся через разные анализаторные входы. В них должен осуществляться уже не только процесс кодирования, но и обязательно – местного декодирования сигнала, формирования местных, в пространстве мозга взаимодополняющих оперативных единиц интегративной деятельности. Этот декодированный сигнал уже не ограничен необходимостью обладать свойствами, приемлемыми для передачи по нервным путям. Его взаимодействие с аналогичным сигналом в остальных задействованных зонах мозга идет по другим законам, а поэтому он может иметь и существенную избыточность. Декодированный сигнал, с одной стороны, изменяет состояние структуры и, с другой – проявляется только на определенном ее функциональном фоне, выявляет свойства структуры, считывающей и шифрующей его в биохимическом коде и вновь «отдающей» его в биоэлектрический код – по требованию.
Паттерны-коды и их элементы – групповые последовательности разрядов – удерживаются или воспроизводятся в том же виде в нейронной популяции на протяжении многих секунд и даже десятков минут в зависимости от характера теста, свойств структуры и ее функционального состояния. Период жизни этих нейродинамических единств, таким образом, ограничен. В первую очередь это относится к групповым последовательностям разрядов с определенными интервалами. Далее может развиться реорганизация импульс ной активности данной нейронной популяции, местная или отражающая общие перестройки в мозгу, и при сохранении того же принципа в паттерне, соответствующем тому же слову, будут возникать и удерживаться на некоторое время те же или уже другие групповые последовательности разрядов, на данный новый отрезок времени также с определенными интервалами. Паттерны и их элементы формируются, удерживаются и распадаются с сохранением в фоновой активности опорных элементов. Популяция нейронов оказывается вновь готовой к деятельности, формированию по тем же принципам новых паттернов и их элементов. Принципиальная возможность этого подчеркивается наличием в мозгу человека нейронов, дифференцированно реагирующих на семантику вербальных тестов (Гоголицын, 1979; Кропотов, 1979б) (рис. 17). В исследовательском плане данные этих авторов определяют поиск мозговых зон с более и менее устойчивыми элементами паттерна и одновременное использование элементов и полного дистрибутивного паттерна при изучении нейрофизиологической организации восприятия, удержания и воспроизведения слов и осуществления мыслительных процессов (Шкурина, 1983).
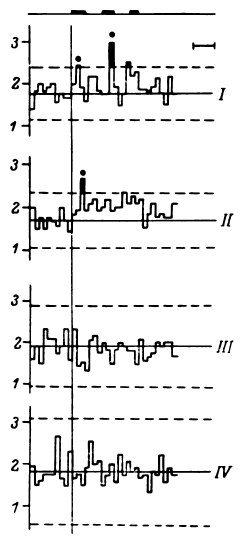
Рис. 17. ПСГ активности одиночного нейрона крючка извилины гиппокампа. Каждая гистограмма накоплена по 12 предъявлениям пар слов, которые обобщались по смыслу пациентом. III – гистограмма, накопленная при предъявлении цифры. Остальные обозначения – как на рис. 16. Видно отсутствие ответа на гистограммах III и IV, а также некоторые различия в положении максимумов на гистограммах I и II (черные кружки)
Подчеркивание динамичности биоэлектрических коррелятов психической деятельности, их неразрывной связи со свойствами мозговых структур, в которых они регистрируются, важно для понимания того, что дальнейшее изучение мозгового кода психических процессов требует не только одновременного использования ряда составляющих комплексного метода изучения мозга, но и прежде всего сопоставления импульсной активности нейронных популяций с медленными электрическими процессами. Оно требует также обязательно сочетанных физиологических и молекулярно-биохимических исследований. Именно в последние годы и этот комплекс становится все более реальным.
На основе использования главным образом показателя структуры импульсного потока были проведены и другие исследования, целью которых являлось получение хотя бы ориентировочного представления о нейродинамике при реализации несколько более сложных психологических тестов. Подробно эти данные приведены нами в первом издании данной книги (1980).
Суть этих работ в изучении способом эталонного поиска того, что развивается в динамике импульсной активности нейронных популяций в условиях, когда испытуемым предъявлялись цветные пейзажи или жанровые картинки в виде слайдов и предлагалось ответить, например, на вопрос о том, к какому сезону года относится данный снимок, или на аналогичный вопрос, детерминирующий принятие решения. Исследование строилось таким образом, что дистрибутивные паттерны – корреляты слов, служащие в дальнейшем эталоном для машинного поиска, выделялись во время однократного исследования продолжительностью в несколько минут и реже – десятков минут с предъявлением этих слов и в том же исследовании реализовалась задача с указанным выше принятием решения. Такие жесткие условия были необходимыми в связи с обнаруженной и описанной динамичностью дистрибутивных паттернов и их элементарных единиц – интервальных последовательностей. Полученные нейрофизиологические данные принципиально не отличались от того, что было приведено ранее в связи с исследованиями, где предъявлялись квазислова, слова, слоги, фонемы и тесты на обобщение слов, имеющих смысловую общность. В импульсной активности нейронных популяций исследованных нами подкорковых структур при рассмотрении слайда и принятия по нему задаваемого решения при эталонном машинном поиске обнаруживались перестройки структуры импульсного потока, выявленные ранее (в том же тесте!) при прослушивании и/или произнесении больным слов, имеющих отношение к содержанию слайда или ответу.
Большинство такого рода работ было проведено нами в 1970-х годах, когда важнейшим вопросом была принципиальная проходимость или, наоборот, непроходимость пути.
В результате этих исследований были получены первые опорные данные о возможности извлечения из динамики импульсной активности коррелятов мыслительных процессов. Большим достоинством этих исследований, которые было бы неправомерно недооценивать, явилось изучение того, что происходит в мозгу в ходе единичного опознания сигнала, однократного принятия решения и т. д., то есть соответствие физиологического исследования тому, как решает свои проблемы мозг человека. В то же самое время динамичность исследованных перестроек импульсной активности, возможность ориентации на них лишь в коротких временны?х промежутках и, таким образом, полное отсутствие рутинно требуемой статистической подтвержденности определили следующий этап работы.
Второй этап исследований. Суть этого, второго, этапа наших работ в той же проблеме изучения нейрофизиологических коррелятов мыслительной деятельности была в решении задачи получения максимально достоверных данных о мозговой нейродинамике при мыслительной деятельности. Это отражало развитие работ, оно было важно для того, чтобы данные обрели необходимую устойчивость и могли быть воспроизведены в любой другой, располагающей соответствующими возможностями лаборатории. Своего рода упрощение в этих работах, однако, определялось тем, что при анализе импульсной активности как бы отступали от того, как реализуется мыслительная деятельность взрослого человека, отступали от изучения нейродинамики, коррелирующей с одиночными тестами. Исследовался так называемый накопленный ответ мозга при предъявлении достаточно большого числа (до 60 и даже более 100) принципиально однотипных проб, соотносимый с фоновой импульсной активностью, сравниваемый в соответствии с фазами теста, его вариациями и характером выполнения проб. Короче говоря, множество различных вопросов решалось при изучении местных процессов способом перистимульных гистограмм (ПСГ). Следует, однако, подчеркнуть, что одновременно, хотя и менее интенсивно, изучалась структура импульсного потока и весьма интенсивно – организация системы обеспечения мыслительной деятельности.
Таким образом, на данном этапе работы основная задача осталась той же – исследование отражения мыслительной деятельности в перестройках импульсной активности нейронных популяций коры и подкорки. Одним из важных отличий от первого этапа была существенная стандартизация тестов, необходимая при исследовании динамики импульсной активности методом ПСГ, обязательным условием которого являлось накопление данных. Испытуемым предъявлялись пробы в виде слайдов в зависимости от задачи с помощью тахистоскопа, светодиодной матрицы или в виде звуковых сигналов. В задачах на опознание в случайном порядке предъявлялись изображения (точнее, силуэты), имеющие смысловое значение, и изображения, одинаковые с первыми по физическим характеристикам, но не имеющие смыслового значения (рис. 18). Изображения, в том числе и цифры, предъявлялись с достаточным для опознания временем или таким образом (по времени), что правильное опознание регистрировалось примерно в 50 % проб.
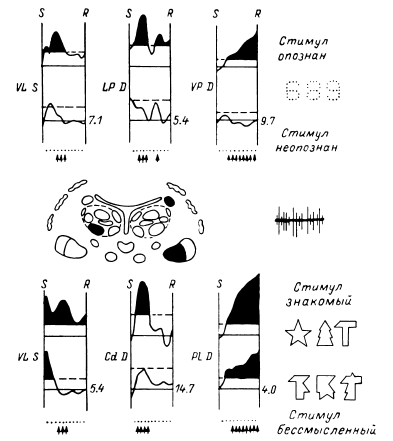
Рис. 18. Примеры пар ПСГ, вычисленных для нейронных популяций в различных образованиях таламуса и стриопаллидарной системы, в пробах с предъявлением цифр на пороге опознания (вверху) и в пробах с предъявлением знакомых и бессмысленных многоугольников (внизу).
S – предъявление стимула; R – начало вербальной реакции больного. VL S – вентролатеральное ядро левого полушария; LP D, VP D, Cd D, Pl D – соответственно заднее латеральное, заднее вентральное, хвостатое и подушечное ядра правого полушария. Точки – отметки времени (бин – 150 мс). Цифры справа внизу каждой ПСГ – средняя частота импульсов в бине. Стрелки указывают на бины, в которых обнаружены значимые различия между средними частотами разрядов для верхней и нижней гистограмм. Непрерывные горизонтальные линии – средние уровни частоты разрядов в фоновом фрагменте, прерывистые– уровни значимости Р < 0.01 отличия от частоты в фоновом фрагменте. Фрагменты гистограммы, выходящие за пределы уровней значимости, отмечены черным цветом
В нейронных популяциях, расположенных в области VA, VL, LP, Cd, CC, Ci, на ПСГ наблюдались коротколатентные компоненты (латентный период до 200 мс), одинаковые при предъявлении и значимых, и незначимых силуэтов изображений. В области VA, VL, LP, Rt, CM, Cd, CC, Ci наблюдались один или два типа длиннолатентных компонентов, причем первый из них (латентный период свыше 250 мс), так же как и коротколатентный, имел форму, близкую к колоколообразной, а второй мог быть характеризован как медленное учащение (или урежение) разрядов нейронов. Эти длиннолатентные компоненты наблюдались при предъявлении значимых стимулов (см. рис. 14). В задачу обследуемого лица в данном тесте входило опознание сигнала, дача ответа, соответствующего изображению в случае значимого сигнала, и ответа «нет», если значение сигнала было неизвестным. Медленное учащение разрядов наблюдалось непосредственно перед словесным ответом и, возможно, характеризовало подготовку к нему.
Таким образом, было обнаружено длиннолатентное учащение разрядов нейронов различных зон мозга, статистически достоверно связанное с опознанием значимых изображений, – отражение в реакции нейронов смыслового элемента проб (Кропотов в кн.: Бехтерева и др., 1985). Для дальнейшего изучения именно этой важнейшей в наших работах позиции сотрудником лаборатории Ю. Д. Кропотовым (1984) проводилось специальное исследование, в котором бессмысленным изображениям-силуэтам придавался определенный смысл. Больной обучался узнавать его как имеющий определенное значение. В импульсной активности нейронных популяций, расположенных в области вентролатерального ядра таламуса, на предъявление ранее незначимого сигнала появились длиннолатентные компоненты (рис. 19). Эти наблюдения еще раз подтвердили, что длиннолатентные компоненты ПГС связаны с отражением смысловых элементов реализуемых проб.
Мне казалось целесообразным здесь достаточно подробно остановиться на этих исследованиях, как существенно подтверждающих своего рода «хрупкие» данные первого этапа. И в то же самое время, как вполне понятно, данная книга не является эпическим повествованием обо всем том действительно огромном материале, который накоплен за годы исследований нейрофизиологии мыслительной деятельности в лаборатории.
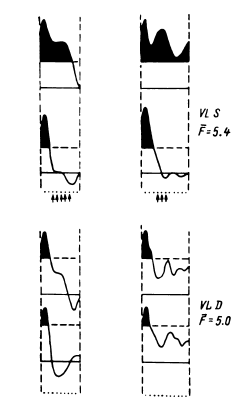
Pис. 19. Сравнительный анализ ПСГ для двух нейронных популяций больного паркинсонизмом, выполнявшего тесты на опознание и обучение.
Слева – пары ПСГ для теста на обучение (верхние гистограммы в каждой паре – суммарный вызванный ответ на первые четыре предъявления стимула, нижние – вызванные ответы на последние четыре предъявления). Справа – пары ПСГ для теста на опознание (верхние гистограммы, в каждой паре – предъявление семантически значимых, нижние – семантически незначимых стимулов). Остальные обозначения – как на рис. 18
Поэтому, например, в отношении исследований с предъявлением сигналов на пороге опознания будут приведены только некоторые данные, а в случае, если читатель захочет подробнее ознакомиться именно с этим вопросом, он может обратиться непосредственно к работам сотрудников нашей лаборатории (Кропотов, 1983а, 1983б; Бехтерева и др., 1985а).
Пробы с предъявлением сигналов на пороге опознания были сконструированы таким образом, что, как указывалось, в 50 % случаях больной не опознавал сигнал или опознавал его ошибочно. В данном случае в прямой зависимости от исследуемой зоны мозга наблюдалась одинаковая активационная (учащение разрядов) или тормозная (урежение разрядов) реакция или избирательная реакция: только на правильное и, что очень важно подчеркнуть, только на неопознание или ошибочное опознание сигнала (рис. 20).
Сопоставимые с этими наблюдениями данные были получены нами впервые вместе с В. Б. Гречиным в 1968 году. При регистрации медленных физиологических процессов мозга – таких, как локальный кровоток и напряжение кислорода, осуществляемой во время выполнения психологических тестов, было обнаружено, что в некоторых зонах хвостатого ядра и таламических образований изменение этих процессов обнаруживалось избирательно при ошибочной реализации теста.
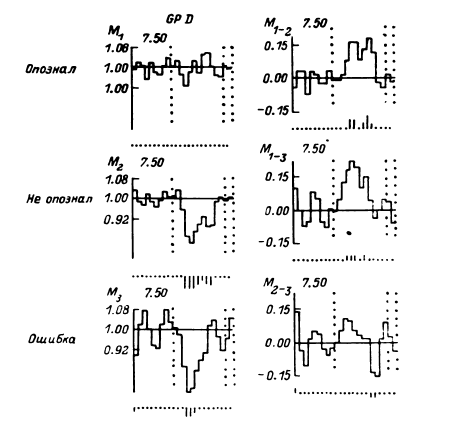
Рис. 20. ПСГ импульсной активности нейронной популяции бледного шара правого полушария для случаев опознания (М1), неопознания (М2) и ошибочного опознания (М3) зрительных стимулов (цифр), предъявляемых на пороге опознания, а также различия между ПСГ (М1–2, М1–3, М2–3).
На M1—М3 по оси ординат – отношение средней частоты разрядов данного бина
На M1—М3 по оси ординат – отношение средней частоты разрядов данного бина к средней частоте фонового фрагмента. На М1–2, М1–3, М2–3 по оси ординат – отношение разности средних частот разрядов соответствующих ПСГ к средней частоте разрядов в данном бине. Под гистограммами – отметки бинов с символическим отражением уровней значимости в длине вертикальных отрезков (Р < 0.05; Р < 0.01; Р < 0.001). Левый вертикальный пунктир – предъявление цифры, два вертикальных пунктира справа – предъявление триггерного стимула, служащего сигналом для вербальной реакции испытуемого. Цифры над гистограммами – среднее число импульсов в бине для фонового фрагмента
Это явление было названо нами детекцией ошибок и соответственно зоны, где оно обнаруживалось, – детекторами ошибок. Конструкция психологических тестов в тех ранних исследованиях не предполагала большого количества ошибок, они были редки – и соответственно еще реже наблюдалась детекция ошибок. Феномен оставался описанным, но трудно было на основании исследования медленных процессов судить о физиологической сущности эффекта. При анализе импульсной активности нейронов в условиях реализации тестов указанной выше конструкции феномен стал «ручным», его можно было наблюдать достаточно большое количество раз в пределах теста для того, чтобы получить статистически достоверные результаты. В зонах мозга, расположенных в области бледного шара, неопознание и ошибочное опознание сигналов соотносилось с фазическим урежением импульсной активности – с реакцией тормозного типа. Следует отметить, что реакции тормозного типа наблюдались в ряде областей мозга и, в частности, в том же бледном шаре и при правильных реализациях теста, и менее селективно – вне зависимости от характера опознания.
Ранее (Бехтерева, Гречин, 1968; Бехтерева, 1971) нами предполагалось, что детекция ошибок является оптимизирующим механизмом мозга, способствующим облегчению последующих мыслительных операций. Местная тормозная реакция в областях мозга, где менее селективные реакции тормозного типа также наблюдаются, может рассматриваться также как элемент оптимизирующих механизмов мозга, хотя вопрос, несомненно, нуждается в дальнейшем уточнении и исследовании. Косвенным свидетельством того, что в данном случае есть некоторые основания думать о торможении тормозных зон, являются наши наблюдения, впервые опубликованные в 1964 году и позднее подтвержденные, о том, что электрическая стимуляция некоторых зон хвостатого ядра и других областей мозга вызывала клинические и поведенческие тормозные реакции.
Вопрос о физиологическом значении явления детекции ошибок интересен в плане изучения общих и частных механизмов мозга и, несомненно, заслуживает дальнейшего исследования. Здесь возможно и важно проследить влияние различных фармакологических средств, а также уточнить, является ли данный феномен специальным атрибутом системы обеспечения мыслительной деятельности (что маловероятно) или это более общий механизм мозга. Заканчивая на сегодня разговор о детекции ошибок, пожалуй, можно сказать, что, каково бы ни было значение этого явления, селективность реакций импульсной активности при психологических тестах в зависимости от качества их выполнения косвенно также свидетельствует в пользу зависимости динамики импульсной активности от содержательных (смысловых) элементов психических процессов. Статистически достоверные данные об изменениях импульсной активности в связи со смысловыми характеристиками теста были получены способом ПСГ и при пробах, требующих реализации относительно более сложных мыслительных операций. Естественно, как и большинство психологических проб, удовлетворяющих условиям многократного повторения в тесте, они были сложнее действительно лишь относительно. Абсолютно они были очень просты, однако, что важно, требовали постоянного внимания и хотя и элементарного, но все же принятия решения о характере ответа.
Одной из основных проб такого типа была проба в форме сравнения двух чисел. Конкретно: больному на экране с помощью светодиодной матрицы предъявлялась цифра, затем буква и снова цифра. Количество светодиодов в каждом знаке было одинаковым. В зависимости от того, какая буква следовала за первой цифрой, надо было просто повторить вторую или сказать, больше или меньше она первой. При этом наблюдались изменения частоты импульсной активности в зависимости от зоны мозга в разные фазы теста (Гоголицын, Кропотов, 1983; рис. 21). Примерно в половине зон мозга наблюдались достоверные различия результатов в зависимости от того, повторялась ли вторая цифра или сравнивалась с первой. Характер различий определялся зоной, с которой отводилась импульсная активность. Цифровые пробы предъявлялись и в звуковой форме, причем различия в зависимости от модальности сигналов были невелики. Различий нейродинамики в зависимости от света, «больше» или «меньше» вторая цифра, не обнаружено. Изменения частоты импульсной активности, отражающие смысловые (мыслительные) операции данного теста, наблюдались в зонах мозга, расположенных в заднем латеральном ядре, срединном центре, переднем и заднем вентральных ядрах, вентролатеральном ядре зрительного бугра, в хвостатом ядре. Следует отметить, что во многих нейронных популяциях, расположенных в области крючка гиппокампальной извилины, вентролатерального ядра зрительного бугра, ретикулярного ядра и в некоторых других структурах, изменений частоты при данном тесте не наблюдалось, что еще раз подчеркивает значимость реакции в тех зонах, где она развивалась.
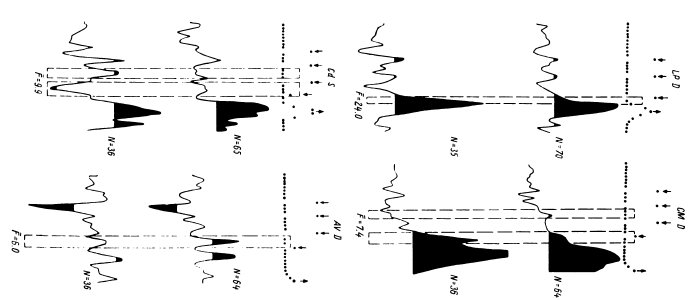
Рис. 21. ПСГ активности четырех нейронных популяций в различных зонах мозга при сравнении по величине двух зрительно предъявленных цифр. Верхние гистограммы – при осуществлении мыслительной операции сравнения цифр, нижние – при назывании второй предъявленной цифры. Точки над гистограммами – отметки времени (бин – 256 мс). Положение точек по вертикали соответствует моментам появления на светодиодном табло первой цифры, буквы, второй цифры, символа, разрешающего вербальный ответ (стрелки вниз), а также отмечает усредненную эпоху звучания ответа испытуемого (стрелка вверх). N – число проб, использованных при накоплении ПСГ. F – средняя частота разрядов на протяжении пробы (имп./бин). Штриховыми линиями выделена область достоверного различия ПСГ. Зачернены участки гистограмм, на протяжении которых наблюдались значимые отклонения частоты разрядов от фонового уровня. LP D – заднее латеральное ядро зрительного бугра правого полушария; CM D – срединный центр зрительного бугра правого полушария; Cd S – хвостатое ядро левого полушария; AV D – переднее вентральное ядро зрительного бугра правого полушария
Во всех этих исследованиях, а также в других аналогичных, несмотря на то что достоверность результатов была очень высока (Р < 0.01 и даже Р < 0.001), вновь проявилась динамичность нейрофизиологических коррелятов мыслительной деятельности. Частотные перестройки импульсной активности, коррелирующие с восприятием физических характеристик тестов и вербальным ответом (в том числе – и подготовкой к нему), были, как правило, устойчивыми в зоне, в которой они первоначально обнаруживались, и воспроизводимыми в последующих исследованиях. Наоборот, перестройки частоты импульсной активности, связанные с опознанием смысла сигнала, с мыслительной операцией сравнения и т. п., были в большинстве зон мозга лабильными; высокодостоверные изменения, наблюдавшиеся сегодня в одной из зон подкорки и коры, могли не наблюдаться в некоторых из этих зон в последующие дни исследования. Более того, при предъявлении в тесте ста проб и последующем анализе импульсной активности в первой и второй половинах теста также обнаруживалось уменьшение достоверности перестроек в некоторых мозговых зонах. Так, например, в тесте на опознание символов, предъявляемых с околопороговыми экспозициями, достоверные изменения в обеих его половинах сохранились в зонах, расположенных в области заднего вентрального ядра зрительного бугра правого полушария, вентролатерального ядра зрительного бугра левого полушария и мозолистого тела левого полушария, и перестали обнаруживаться в зонах в области хвостатого ядра левого полушария и заднего латерального ядра правого полушария (рис. 22). Это явление еще раз подтвердило выдвинутые ранее представления о наличии в мозговых системах обеспечения мыслительной деятельности не только постоянных, жестких звеньев, но и большого количества звеньев переменных – гибких. Динамичность мозговых систем обеспечения мыслительной деятельности является, по-видимому, одним из важных механизмов информационной емкости мозга (Бехтерева и др., 1983).
Кроме сказанного в связи с этими исследованиями важно подчеркнуть еще раз по крайней мере две позиции. Во-первых, изменения, которые достоверно соотносились именно с мыслительной деятельностью, наблюдались далеко не во всех зонах мозга. И, во-вторых, перестройки импульсной активности, связанные с восприятием физических характеристик сигнала и вербальным ответом, были существенно более воспроизводимыми, чем те, которые были связаны с отражением в импульсной активности мыслительных характеристик теста.
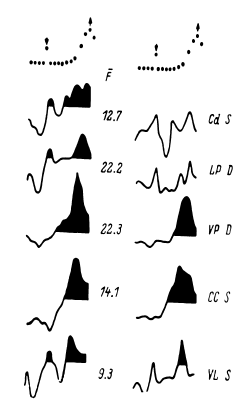
Рис. 22. ПСГ активности пяти нейронных популяций в различных подкорковых структурах мозга в тестах на опознание символов, предъявляемых с околопороговыми экспозициями (300 мкс).
Левая колонка – усреднение первых 49 проб теста, правая – последних 49 проб. Бин – 256 мс. СС S – мозолистое тело левого полушария. Остальные обозначения – как на рис. 18 и 21
Первая позиция вновь подтверждает положение о том, что обеспечение мыслительной деятельности осуществляется мозговой системой, а не мозгом «в целом». Вторая – наглядно демонстрирует разницу мозговых систем обеспечения функций, наличие в мозговой системе обеспечения, например, двигательной деятельности (в данном случае – речедвигательной), значительно большего числа жестких элементов. Таким образом, в исследованиях нейронных коррелятов мыслительных процессов было подтверждено высказанное ранее предположение о связи фактора гибкости в мозговых системах со сложностью (нестереотипностью) обеспечиваемой деятельности (Бехтерева, 1971, 1980б).
Данный этап исследований по отношению к первому может быть обозначен не как второй этап, а точнее, как второй виток спирали в изучении проблемы, так как, по существу, основная проблема сохранялась, однако основной задачей оказывалось обнаружение уже не просто коррелятов мыслительных процессов, но, как указывалось, – статистически достоверных коррелятов. Исследования были прежде всего сосредоточены на изучении перестроек частоты импульсной активности, однако рассматривались с тех же методологических позиций и перестройки структуры динамики импульсной активности. Проводились поиски и изучение не только конкретных последовательностей интервалов, но более общей связи (и прежде всего, ее наличия) между реализацией мыслительной деятельности и изменениями структуры импульсной активности нейронных популяций. Использовался показатель нормированной гистограммы плотности распределения межимпульсных интервалов, то есть принципиально – отношения исследуемых интервалов к средней величине межепайкового интервала. A priori можно было предполагать, что если в этих условиях и выявятся какие-то изменения, то они явятся производным перестройки частоты импульсного потока. Однако, как показали работы Н. Г. Шкуриной (1984), наблюдался не этот один, а три (все три!) теоретически возможных варианта соотношений частоты и структуры импульсной активности. Регистрировались достоверные изменения частоты без достоверных изменений структуры; наблюдалось сочетанное изменение обоих показателей; и наконец, что особенно интересно, изменения структуры потока, как и в предыдущих двух ситуациях, соотносимые с фазами психологического теста, могли наблюдаться изолированно, без существенного увеличения частоты импульсной активности (рис. 23, а, б, в).
Анализировалась импульсная активность 5 популяций вентролатерального ядра зрительного бугра, 2 – бледного шара, 3 – хвостатого ядра и активность, отводимая из мозолистого тела. Следует отметить, что различные соотношения динамики частоты импульсной активности могли наблюдаться как в разных популяциях и, таким образом, в зависимости от зоны мозга, так и в одной и той же популяции, но уже в зависимости от характера теста. Эти материалы подтвердили на новом уровне, что изменения структуры импульсного потока были правомерно отнесены к нейрофизиологическим коррелятам мыслительной деятельности. Эти же данные показали, что перестройки структуры импульсной активности могут иметь в этом плане самостоятельное значение, а отсюда допустимо рассматривать перестройки частоты и структуры импульсной активности как имеющие взаимодополняющее значение. Доказательства значения перестроек структуры импульсной активности как одного из нейрофизиологических коррелятов мыслительной деятельности были получены также в исследованиях С. В. Медведева и Н. И. Цыганкова (1985), проведенных способом авторегрессионного анализа (рис. 24, 25).
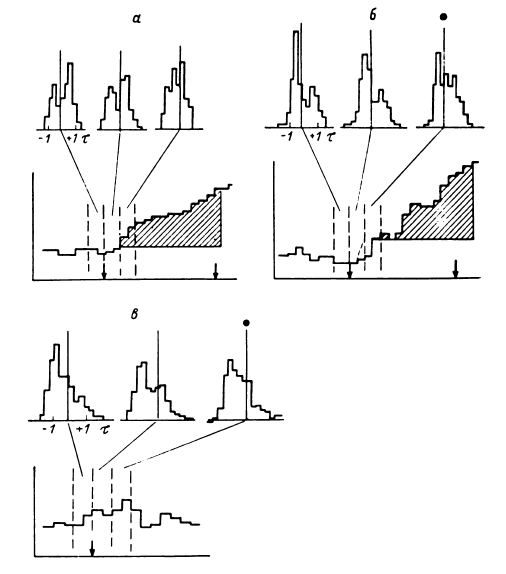
Рис. 23. Нормированные гистограммы плотности распределения межимпульсных интервалов и ПСГ активности нейронной популяции в бледном шаре правого полушария при выполнении тестов на опознание (а, б) и запоминание (в).
Вверху – нормированные гистограммы распределения межимпульсных интервалов, соответствующие выбранным эпохам анализа (ограничены вертикальными штриховыми линиями) (по оси абсцисс – величина межимпульсного интервала, ед. стандартного отклонения – ?; по оси ординат – число межимпульсных интервалов, имеющих данную величину); внизу – ПСГ (по оси абсцисс – время: бин – 150 мс; по оси ординат – число импульсов в бине, усредненное по заданному числу проб: а – 53 пробы, б – 53, в – 107). Заштрихована область высокодостоверного ответа на ПСГ. Стрелками указаны моменты предъявления стимулов и начала ответов испытуемого. Точками отмечены гистограммы, значимо отличающиеся по форме от гистограмм, накопленных по двум другим эпохам анализа
С помощью приема линейной авторегрессионной функции 4-го порядка (вычислялись первые 4 коэффициента) исследовались нейронные популяции, расположенные в стриопаллидарной системе, лимбических образованиях и различных областях коры. Прием позволил выделить из популяций такие субпопуляции, в которых импульсная активность была структурирована. При этом обнаружены своего рода сверхстабильные ансамбли, зависимые от условий исследования. Этот способ анализа оказался выгодным для выделения из нейронной популяции ансамбля, группы нейронов, образующих рабочую единицу при выполнении какой-либо деятельности.
Рис. 24. Динамика структурных коэффициентов bj во время выполнения корректурного теста (мозолистое тело). Вертикальными линиями разделены последовательные повторения задания. Штриховой линией показан уровень bj=0.1. Как видно, импульсная активность структурирована на уровне b3. Динамика b2 отражает временную структуру теста
Рис. 25. Зависимость нормированного структурного коэффициента Y1от уровня амплитудной дискриминации в двух исследованиях (а, б) у одного больного.
По оси абсцисс – среднее значение текущей частоты при различных уровнях амплитудной дискриминации, Гц. Штриховыми линиями ограничен уровень достоверности Р = 0.05. Видна зависимость структуры потока от уровня дискриминации, то есть от размера регистрируемой субпопуляции. Обозначения структур мозга – как на рис. 18 и 21
В исследовании мозговых систем обеспечения деятельности безусловно необходимо изучение местных событий в различных зонах мозга и динамики этих местных событий. На этой основе появляются известные основания для суждения о свойствах мозговой системы обеспечения деятельности. Однако исследование собственно системы должно включать и исследование взаимоотношений между активными зонами мозга – предположительно – звеньями системы.
Такого рода исследования проводились приемом корреляционного анализа, с изучением соотношений в разных зонах мозга изменений частоты импульсной активности и методом анализа динамики совпадений разрядов в микроинтервалы времени в различных зонах мозга.
Исследование соотношений частоты разрядов в различных зонах мозга, активных при осуществлении психологических тестов наряду с самим фактом наличия корреляции между этими зонами показало отсутствие обязательной зависимости тесноты связи между различными зонами мозга от местных изменений частоты импульсной активности (рис. 26). Это позволило рассматривать и третий показатель – соотношение активности в звеньях мозговых систем– также как дополнительный мозговой механизм в обеспечении мыслительных процессов. Таким образом, нами рассматриваются как взаимодополняющие механизмы в нейрофизиологическом кодировании мыслительных процессов изменения частоты, структуры импульсной активности и взаимоотношений этой активности в звеньях мозговой системы.
Как видно из приведенного рисунка (см. рис. 26), по-видимому, следует в прямое развитие сказанного подчеркнуть, что уменьшение выраженности изменений частоты импульсной активности, вплоть до исчезновения достоверных сдвигов, еще не может расцениваться как выключение зон из обеспечения деятельности. И наоборот, простой прием корреляционного анализа может использоваться для уточнения степени: действительно ли зона мозга перестала участвовать в обеспечении деятельности как звено системы.
Рис. 26. ПСГ импульсной активности, зарегистрированной в лучистом венце (левая колонка), премоторной коре правого полушария (правая колонка), и динамика коэффициента корреляции между текущей частотой разрядов этих популяций на протяжении пробы (средняя колонка).
Верхний ряд гистограмм – данные для проб первой половины теста на сравнение по величине двух цифр (49 проб); нижний ряд – данные для проб второй половины теста (48 проб). F – частота, F – средняя частота в бине, R – коэффициент корреляции. Остальные обозначения – как на рис. 18
Исследование динамики совпадений разрядов в зонах мозга в микроинтервалы времени проводилось по принципиальной схеме, предложенной Герштейном (Gerstein et al., 1978) и адаптированной Е. М. Кроль и С. В. Медведевым (1982). Осуществлялось исследование достоверного увеличения числа совпадений импульсной активности в различных популяциях, статистический анализ неслучайного числа совпадений в мозговых зонах, расположенных в области мозговых образований.
Основным психологическим тестом, используемым на данном этапе для исследования соотношений импульсной активности в различных зонах мозга, был так называемый корректурный тест. Перед больным на экране проецировалась таблица из 14 строчек колец с разрывами, имеющими 8 различных ориентации. В некоторые строчки были включены и полностью замкнутые кольца. Предлагалось считать кольца с определенной ориентацией разрыва или отыскивать замкнутое кольцо, которое, кстати, не обязательно присутствовало на слайде. Каждый вид пробы повторялся по 8 раз. Слайд предъявлялся несколько быстрее, чем можно было полностью просмотреть все строки таблицы (около 150 с). Такого рода монотонный длительный тест использовался с целью активации и дальнейшего поддержания в мозгу в активном состоянии определенной системы. Предполагалось, что в этих условиях взаимодействия между звеньями этой системы могут быть изучены наилучшим образом. Исследования С. В. Медведева и сотрудников (Бехтерева и др., 1984; Медведев, Белов, 1986) подтвердили эффективность данной методики и показали, что количество совпадений импульсных разрядов нейронов между дистантными звеньями системы в большей мере определялось характером выполняемой деятельности (рис. 27, 28). Было показано также, что существенное увеличение или, наоборот, уменьшение числа совпадений разрядов по отношению к фоновому уровню может наблюдаться в процессе реализации пробы между некоторыми мозговыми зонами в очень короткие интервалы времени, в том числе в интервалы длительностью 1–2 мс. Эти факты поставили задачу специального рассмотрения физиологических условий такого рода синхронизации разрядов, так как в этом случае трудно было понять эту синхронизацию без введения представлений о пейсмекерном механизме (см. выше представления о динамическом пейсмекере) или о возможности скоростного взаимодействия между дистантными зонами мозга.
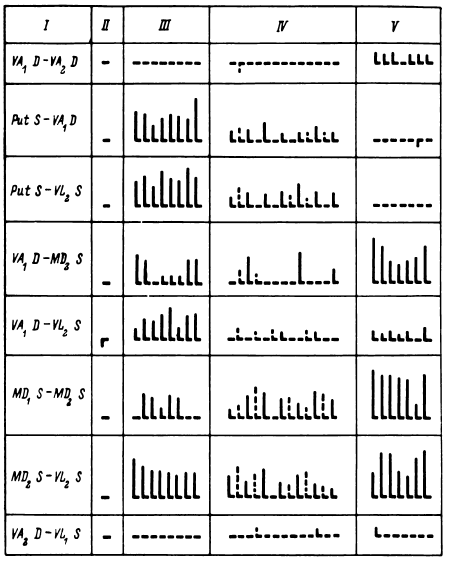
Рис. 27. Динамика связанности между дистантно расположенными нейронными популяциями. I – нейронные участки мозговых структур; II – фон (отсутствие целенаправленной деятельности); III – корректурные пробы со счетом; IV – корректурные пробы со счетом, перемежающиеся сюжетом; V – корректурные пробы без счета. По оси абсцисс штрихами обозначены последовательные пробы; по оси ординат отложена величина квантили (для Р < 0.975). Калибровка по вертикали (длина штриха в колонке IV); квантиль равна 2.0. Сплошные вертикальные линии – корректурные пробы, штриховые – сюжетные пробы. Характеристики связей: T=1 с, ?=l мс
Рис. 28. Паттерны стабильной связанности нейронных популяций.
А—Д – больные, черными точками указаны исследованные популяции. Жирными линиями соединены стабильно связанные популяции
Эту последнюю возможность нельзя исключить полностью в связи с обнаруженным в специальных условиях наблюдения очень резким падением импеданса между двумя звеньями системы, в этих специальных условиях связанной с обеспечением движения (Бехтерева и др., 1980). В связи с трудностью исследования этого вопроса у человека, вероятно, следует организовать целенаправленное его изучение в эксперименте на животных, тем более что в данной ситуации совершенно не обязательно проводить исследования именно системы обеспечения мыслительных процессов.
В проведенных работах достаточно неожиданно для тех, кто конструировал тесты, было обнаружено, что длительное осуществление однообразной деятельности совсем не обязательно коррелирует с однообразием ситуация в мозгу. Было показано, что соотношения в системе обеспечения мыслительной деятельности перестраиваются независимо от длительности монотонного теста на протяжении 0.5–1 с. Иными словами, повышенная синхронизация разрядов нейронов, улавливаемая методом совпадений, наблюдается попеременно между различными активными зонами мозга. Высокая синхронизация активности нейронов в каждый данный микроинтервал времени может наблюдаться далеко не между всеми активными точками, что может быть проиллюстрировано рис. 29.
Следует подчеркнуть не только динамичность связей, но и возможность как бы независимых связей между парами нейронных популяций. Нетранзитивность связей была обнаружена независимо от расстояния между исследованными зонами. Кроме того, по аналогии с местными явлениями были обнаружены и связи разной степени жесткости. Таким образом, и в этом случае была обнаружена динамичность мозгового обеспечения мыслительной деятельности, в данном случае в форме динамической реорганизации системы на протяжении монотонной деятельности. Если в научном исследовании позволены образные выражения (а почему бы и нет?), создается впечатление, что мозг как бы «борется» против монотонности. Динамичность, которую мы наблюдали в местных процессах, является, по-видимому, важным общим принципом работы мозга.
Рис. 29. Схематическое представление элементов системы обеспечения монотонной деятельности (корректуры) с сохранением функциональной структуры действия.
Вверху – корректурные пробы со счетом, внизу – без счета. Характеристики связей: Т=1с, ?=1мс
Что же касается самого методического приема – исследования совпадений разрядов, то он оказался адекватным именно для изучения соотношений активности в небольшие интервалы времени, давал своего рода «мгновенные снимки» ситуации в системе обеспечения мыслительных процессов. При больших бинах (больше, чем 1–3 мс) перестройки, связанные с заданной деятельностью, не выявлялись (Бехтерева и др., 1984). Отсюда, по-видимому, для исследования взаимоотношения местных процессов в звеньях мозговых систем (активных при какой-то деятельности зонах мозга) целесообразно использовать не только один, но целый ряд приемов, в том числе и таких, которые не только позволяют выявить соотношение активности в различных зонах мозга, но и вскрыть различные механизмы организации системы.
Третий этап исследований может быть характеризован как попытка минимизации дефектов методов анализа, связанных с накоплением и таким образом усреднением данных, то есть как бы вновь, на новом витке спирали, попытаться использовать для изучения мозга приемы, более адекватные механизмам мозга. В этом плане предложен и используется компонентный анализ (Гоголицын, Пахомов, 1984, 1985), резервы которого, повидимому, еще не полностью раскрыты. Принцип метода – в первоначальном сохранении максимума информации о динамике местных перестроек в мозгу. Проводится количественное описание импульсной активности в рамках более гибких представлений о паттернах текущей частоты. Это достигается представлением физиологических мозговых событий по каждой пробе в виде диаграмм таким образом, что по оси абсцисс отложено время развития перестроек частоты (ее учащения или урежения), а по оси ординат – выраженность этих перестроек в форме амплитудного показателя (количество импульсов в бине).
Последовательное автоматизированное наложение на одно изображение (диаграмму) данных по множеству проб далее сопровождается выделением кластеров, то есть выделением областей максимумов сгущений точек (рис. 30). Сравнение получаемых с помощью данного приема результатов с ПСГ показало, что ПСГ, во-первых, действительно может отражать события, достаточно близкие к тому, что происходит в импульсной активности нейронов по ходу всего теста, во-вторых, может отражать их приблизительно и, наконец, за низкоамплитудной ПСГ могут скрываться существенные отклонения импульсной активности от фона, но различные по направленности, амплитуде и времени развития на разных фазах теста в каждой пробе (рис. 31). Применение компонентного анализа позволило, таким образом, продемонстрировать, из каких реальных физиологических событий, происходящих при однократных воздействиях стимула, складывается усредненная картина ПСГ.
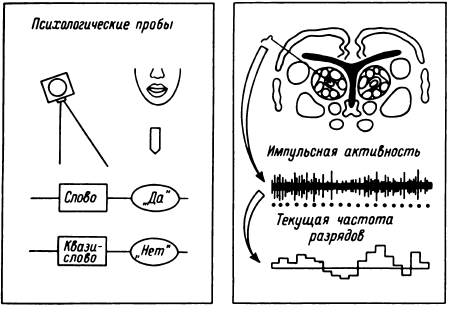
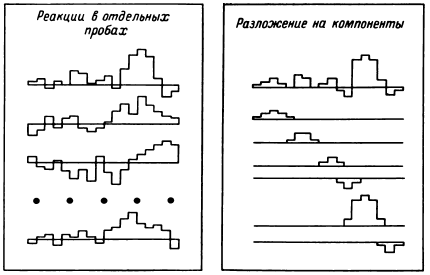
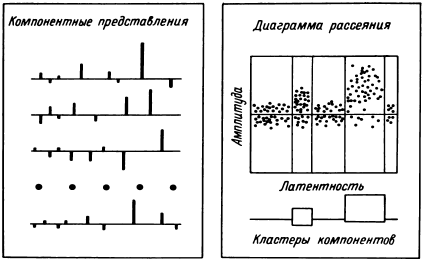
Рис. 30. Схематическое изображение основных этапов компонентного анализа импульсной активности нейронов.
Импульсная активность регистрируется при выполнении психологических тестов, состоящих из однотипных проб. Исходным материалом для анализа являются кривые, описывающие изменения частоты разрядов нейронов в отдельных пробах. Каждая такая кривая с помощью специального алгоритма раскладывается на компоненты, то есть представляется в виде суммы колоколообразных пиков, характеризуемых латентностью, амплитудой и длительностью. Это позволяет получить компонентные представления реакций в отдельных пробах. Используя латентные периоды и амплитуды всех компонентов, можно построить диаграмму рассеяния, наглядно иллюстрирующую степень вариабельности реакций в отдельных пробах на протяжении теста.
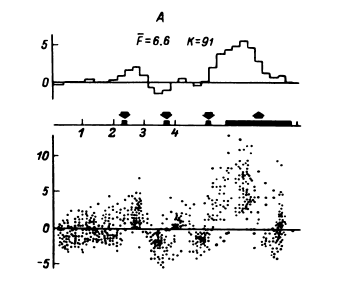
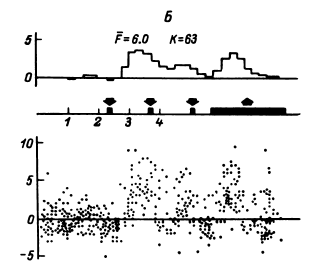
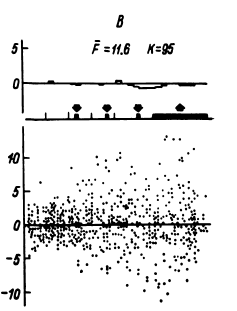
Рис. 31. ПСГ и диаграммы рассеяния «латентный период—амплитуда» активности нейронных популяций в хвостатом ядре левого полушария (А), мозолистом теле левого полушария (Б) и премоторной коре правого полушария (В) в тесте на сравнение цифр по величине.
Под ПСГ на оси времени (цена деления – 1 с) черными прямоугольниками отмечены эпохи предъявления (стрелки вниз) символов (цифра, буква, цифра) и звучание ответа испытуемого (стрелка вверх). По оси ординат ПСГ – отклонение частоты разрядов (имп./бин) от среднего уровня в фоне. Бин ПСГ – 256 мс. F – средний уровень частоты разрядов (имп./бин) на протяжении фонового фрагмента (от начала пробы до момента предъявления первой цифры). К – число проб. ПСГ и диаграмма рассеяния построены в одинаковом масштабе.
Вторым важным направлением данного этапа исследований является сравнительная характеристика функциональных свойств различных мозговых зон. С этой целью анализ динамики импульсной активности ряда нейронных популяций подкорки и коры (чаще всего способом ПСГ) осуществляется при предъявлении больному не какого-то одного, а совокупности, батареи, тестов. В этом случае в полном соответствии с более ранними данными о полифункциональности нейронных популяций (Бехтерева, 1971) были обнаружены разные реакции импульсной активности нейронов коры и подкорковых зон мозга на психологические тесты с различием конкретных заданий в пробах (Кропотов, Пономарев, 1985). Так, на рис. 31 представлены ПСГ, построенные на основе анализа импульсной активности нейронных популяций, расположенных в различных зонах коры и подкорковых структур. Как видно из рис. 32, ПСГ могли различаться при разных психологических пробах очень значительно. На данном рисунке наибольшая изменчивость в зависимости от теста наблюдалась в отведениях одной из популяций бледного шара, вентролатерального ядра зрительного бугра и теменной коры (в области поля 40, по Бродману).
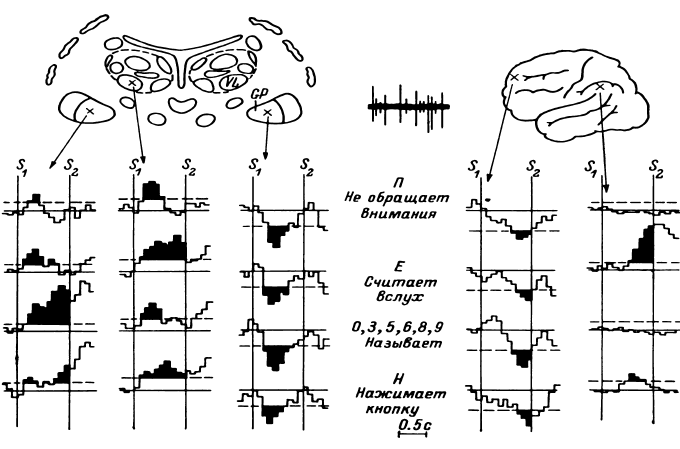
Рис. 32. Примеры ПСГ, вычисленных для различных нейронных популяций корковых и подкорковых структур, в тесте с предъявлением зрительных стимулов, вызывающих различные ответные реакции больных.
S1 – предъявление стимула, определяющего ответную реакцию; S2 – предъявление паттерна шахматной доски, служащего сигналом для ответной реакции. Реакции больного в ответ на предъявление стимулов следующие: буква П – игнорируется, буква Е – подсчитывается текущее число таких стимулов, цифры называются вслух, буква Н – больной нажимает кнопку. Стрелками указаны структуры, в которых регистрировалась импульсная активность. Зачернены участки ПСГ, соответствующие уровню значимости Р < 0.01
Аналогичное явление наблюдалось при предъявлении проб, адресуемых к разным сенсорным входам (зрительным и слуховым) и с речевым ответом или без речевого ответа (рис. 33).
Из приведенных данных можно с учетом динамичности нейронных коррелятов импульсной активности сделать следующие осторожные заключения. В нейронных популяциях, расположенных в области бледного шара, обнаружены реакции как активирующего, так и тормозного типа. Эти реакции были сходными и различными при разных психологических пробах. При этом селективность реакций тормозного типа, если они наблюдаются в отведениях из бледного шара и передних отделов лобной коры, невелика. В нейронных популяциях, расположенных в области вентролатерального ядра зрительного бугра, в области теменной коры, в отдельных популяциях бледного шара, обнаруживаются также реакции активирующего типа, однако они более или менее высокоселективны, различны при разных пробах.
Сравнение между собой реакций тормозного типа (см. выше) показывает, что урежение частоты импульсной активности, независимо от вида мыслительной деятельности, наблюдалось раньше в подкорке (бледный шар), чем в коре (передние отделы лобной коры).
Эти данные, однако, должны расцениваться пока как вероятность появления такого рода нейродинамики. Только длительные повторные исследования позволят уточнить, насколько именно вероятна та или другая из описанных ситуаций. И в то же время уже можно говорить о более или менее селективной реакции на психологические пробы в различных нейронных популяциях.
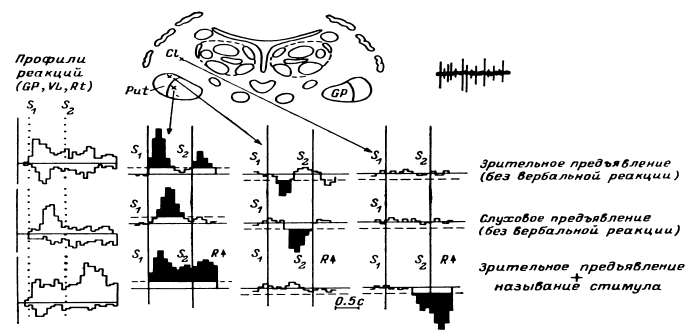
Рис. 33. Реакции нейронов мозга человека в ответ на предъявление зрительных и слуховых стимулов.
Профили реакции всех обследованных нейронных популяций (слева) и ПСГ, вычисленные для отдельных нейронных популяций подкорковых образований мозга в ответ на зрительное (верхний ряд), слуховое (средний ряд) предъявления цифр в отсутствие вербальной реакции и в случае, когда человек называет цифру вслух (нижний ряд). По оси ординат на профилях реакций – число нейронных популяций, в которых были обнаружены значимые (Р < 0.05) отличия частоты разряда в бине от средней фоновой частоты, на ПСГ – средняя частота разряда в бине; по оси абсцисс – время в бинах (бин – 100 мс). На гистограммах: S1 —предъявление информационного стимула (цифры); S2 – предъявление триггерного стимула, служащего сигналом для вербальной реакции испытуемого; R – название стимула. Остальные обозначения – как на рис. 32
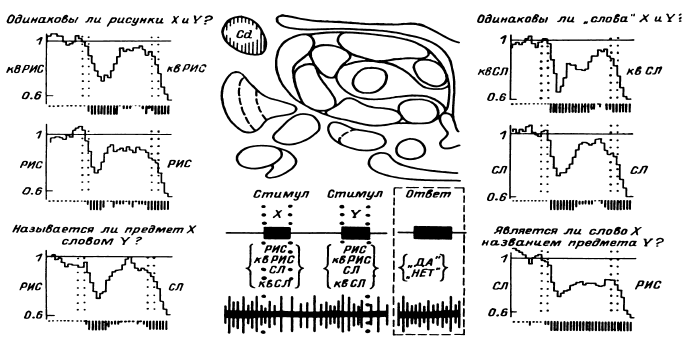
Рис. 34. ПСГ импульсной активности нейронной популяции в хвостатом ядре левого полушария при выполнении серии тестов, связанных со сравнением слов, квазислов, рисунков известных и неизвестных объектов.
Схематическое изображение структуры пробы дано в середине нижней части рисунка. В любой пробе предъявляются два стимула, каждый из которых может быть рисунком известного (РИС) или неизвестного (кв РИС) объекта, словом (СЛ) или квазисловом (кв СЛ). В зависимости от даваемой перед началом теста инструкции испытуемый в каждой пробе отвечает «Да» или «Нет» с учетом особенностей предъявленных стимулов
Бо?льшую или меньшую селективность реакций нейронных популяций удалось наблюдать и при предъявлении совокупностей тестов с опознанием слов, сравнением слов и квазислов, слов с соответствующими и не соответствующими им изображениями, сравнением изображений, имеющих и не имеющих смыслового значения, и т. п. Иллюстрацией сказанного может служить рис. 34 (Гоголицын и др., 1987б).
Особую осторожность (опять с учетом динамичности нейронных коррелятов мышления) следует проявлять в этих исследованиях при оценке негативных эффектов – когда нет значимых изменений импульсной активности при психологических пробах. При наличии перестроек на основе одного-двух тестов правильнее говорить, что данная зона, являясь звеном системы, может более или менее постоянно участвовать в обеспечении данного вида деятельности. При отсутствии перестроек для того, чтобы сказать, что данная зона мозга не может участвовать в обеспечении реализуемой деятельности, безусловно, необходимы повторные исследования (особенно это относится к случаям с высокой селективностью реакций). С учетом этих положений, может быть, следует осторожнее относиться к данным, полученным в результате электрической стимуляции открытого мозга (Ojemann, 1983), на основе которых описывается высокая функциональная селективность в обеспечении мыслительных и преимущественно вербально-мнестических процессов.
Исследования импульсной активности нейронных популяций подтвердили многие механизмы мозга, показанные или предсказанные нами ранее. Эти работы безусловно явились и существенно новой ступенью в изучении различных мозговых механизмов. Это, в частности, относится к динамичности мозговых коррелятов мыслительной деятельности, которая была, во-первых, гораздо полнее изучена, доказана и, во-вторых, рассмотрена более полно в плане внутренних механизмов, определяющих динамичность нейрофизиологических коррелятов мыслительных процессов.
Ранее нами (Бехтерева и др. 1970) было показано, как могут изменяться свойства зоны мозга в зависимости от состояния различных видов биохимической медиации. Позднее В. А. Илюхиной (1977) была обнаружена зависимость свойств мозговых зон от уровня так называемого постоянного потенциала, наиболее стабильной составляющей СМФП – омега-потенциала (по: Илюхина, 1982б). При одновременном отведении и исследовании импульсной активности нейронных популяций менее стабильных СМФП была обнаружена зависимость функциональной характеристики зоны мозга и от этого показателя.
В исследованиях С. В. Медведева и сотрудников (Медведев, Белов, 1985, 1986; Медведев и др., 1987) регистрировалась динамика импульсной активности нейронов (ИАН) при одновременном отведении с той же точки мозга, с того же электрода (что определялось возможностями полиэлектронейрографа, – Данько, Каминский, 1982) СМФП в декасекундном диапазоне (тау-волн, по В. А. Илюхиной). Больному предъявлялись психологические пробы на обобщение зрительного стимула.
В зонах мозга, расположенных в области различных ядер таламуса, стриопаллидарной системы была обнаружена зависимость динамики ИАН по ходу реализации психологического теста от фазовых характеристик тау-волн в той же зоне (рис. 35). Особенно интересной была в этом случае тонкая модуляция функциональных свойств мозговых зон.
Как представлено в работах В. А. Илюхиной (1977, 1986), при изменениях наиболее стабильной составляющей СМФП наблюдались перестройки функциональных свойств зон мозга, которые можно было характеризовать типом «да–нет», то есть свойство либо исчезало, либо появлялось. При изменениях фазы тауволн могло наблюдаться уменьшение или увеличение изменений ИАН в различные фазы психологического теста. Так, например, в зоне, расположенной в области заднего вентроорального ядра таламуса, при реализации психологического теста на восходящей и частично нисходящей фазе тау-волны существенные изменения ИАН обнаруживались в фазе принятия решения. В той же зоне при реализации того же теста на нисходящей–восходящей (рис. 35) фазе тау-волны изменения частоты ИАН в фазу принятия решения отсутствовали.
При предъявлении в тесте двух абсолютно одинаковых сигналов, из которых только первый нес информационную нагрузку, а второй был в контексте теста незначим, на восходящей– нисходящей фазе тау-волны обнаруживалась реакция только на первый из двух сигналов. А при предъявлении их на нисходящей– восходящей фазе селективность реакции зоны мозга исчезала – зона реагировала одинаково на оба сигнала.
Рис. 35. Зависимость динамики импульсной активности нейронов от фазовых характеристик тау-волн тех же зон мозга при мыслительной деятельности.
А – результаты усреднения СМФП в декасекундном диапазоне по положительным (б, г, е) и отрицательным (а, в, д) группам знакового ключа; Б – ПСГ текущей частоты импульсной активности нейрона: а, б, д, е – смысловые группы; в, г – «пустышки»; а–г – регистрация в области заднего вентроорального ядра таламуса (V. о. р); д, е – регистрация в области заднего дорсоорального ядра таламуса (D. о, р). Вертикальными штриховыми линиями обозначены моменты предъявления I (S1), II (S2) стимулов и сигнала, разрешающего речевой ответ больному (SR)
Такого рода модулирующее влияние тау-волн обнаружено не во всех исследованных зонах и не во всех зонах с одинаковой выраженностью. Обнаруженная зависимость должна изучаться и, может быть, через некоторое время удастся уточнить, какова степень общности данного модулирующего механизма мозга. В данном случае именно выявление его в каких-то зонах мозга должно расцениваться как импульс к дальнейшему поиску, а не как отсутствие их и, следовательно, как повод к «закрытию проблемы».
Исследования нейрофизиологических механизмов мыслительной деятельности, несомненно, целесообразно продолжить, чтобы шаг за шагом получать все более полные характеристики вклада различных мозговых зон коры и глубоких структур мозга в обеспечение мыслительной деятельности. Одновременно важно дальнейшее выяснение механизмов мозга, их общего и частного значения. Мы очень ждем от работающих вместе с нами представителей точных наук методов исследования, все более адекватных тому, как мозг взрослого человека решает простые, сложные и сложнейшие задачи. Надо также надеяться, что в самом ближайшем будущем изучение местных и системных процессов в мозгу человека сольется в единое целое. Вместе с психологами физиологи должны попытаться понять мозговые механизмы не только мыслительной деятельности, обеспечивающей ежедневную жизнь человека, но и тех процессов, которые служат прорыву в неизвестное, то есть механизмы творчества. Однако какими бы интригующими ни были результаты будущего, они не явятся поводом для третьего издания данной книги. В задачи книги входило наряду с детализацией ряда данных прежде всего изложение общих подходов к изучению здорового и больного мозга и общих принципов его работы.






