Репликация ДНК
Процесс самоудвоения молекулы ДНК, обеспечивающий точное копирование генетической информации, называется репликацией или удвоением молекул ДНК.
Открытию механизма репликации предшествовали многочисленные эксперименты по синтезу ДНК. Модель молекулы ДНК, предложенная Джеймсом Уотсоном и Френсисом Криком, породила целую серию экспериментов по выяснению механизма ее репликации. Наиболее убедительные данные были представлены в 1958 г. учеными М. Мезелсоном и Ф. Сталем. В процессе эксперимента были выдвинуты три гипотезы.
1. Консервативная репликация. Двухцепочечная молекула ДНК служит матрицей для синтеза полностью новой молекулы ДНК, т. е. новая ДНК является полной копией исходной.
2. Полуконсервативная репликация. На каждой цепи исходной молекулы ДНК синтезируется вторая недостающая цепь по принципу комплементарности. Каждая новая ДНК состоит из одной исходной и одной новой цепи ДНК.
3. Фрагментарная репликация. Молекула ДНК распадается на короткие фрагменты, которые используются в качестве матрицы для достройки недостающих фрагментов новых молекул ДНК.
В качестве объекта эксперимента была использована кишечная палочка, которая содержит в клетке всего одну молекулу ДНК. Для выяснения правильной версии бактерии выращивались на питательной среде, содержащей радиоактивный изотоп азота 15N. Через несколько поколений ДНК всех бактерий содержали изотоп 15N (рис. 24).
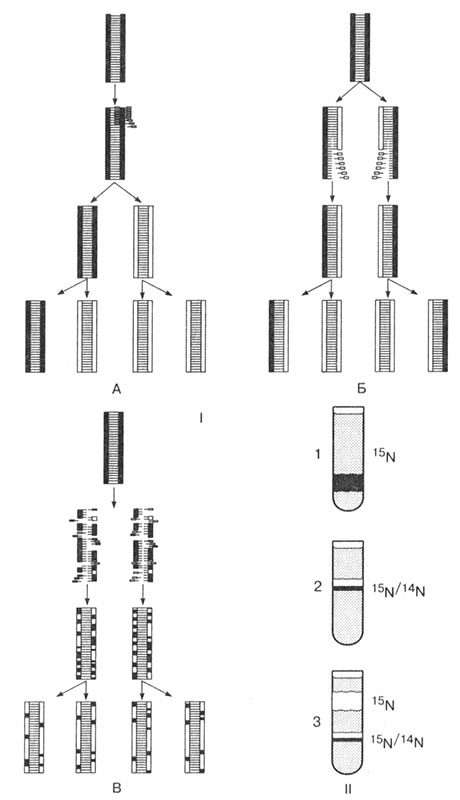
Puc. 24. Гипотезы процесса репликации ДНК: 1. А — консервативная; Б — полуконсервативная; В — фрагментарная. II. Результат центрифугирования молекулы ДНК бактерий
Методом центрифугирования эти ДНК были выделены из клеток в виде отдельной фракции, что доказывало, что ДНК всех бактерий имеют одинаковый изотопный состав. Далее бактерии, содержащие в ДНК только изотоп 15N, были перенесены на питательную среду с обычным изотопом азота 14N. Центрифугирование ДНК нового поколения бактерий показало, что эти молекулы содержали азот 14N и 15N, но их невозможно было разделить по массе на фракции. Следовательно, оба изотопа находились в одной молекуле ДНК. Первая версия о консервативном способе репликации ДНК была отвергнута, так как отсутствовали отдельные фракции ДНК с 14N и ДНК с 15N.
Для проверки двух оставшихся версий были исследованы ДНК третьего поколения бактерий. Их удалось четко разделить на две фракции: ДНК (14N) и ДНК (14N, 15N). Таким образом, отпала третья версия о фрагментарной репликации ДНК. Эксперимент позволил установить, что ДНК синтезируется полуконсервативным способом.
В основе репликации ДНК лежат следующие принципы.
Комплементарность. Каждая цепь молекулы ДНК содержит последовательность нуклеотидов, в точности комплементарную последовательности нуклеотидов на другой цепи. Следовательно, новые молекулы несут одну и ту же генетическую информацию: цепь А является шаблоном для синтеза цепи А', а цепь А' — шаблоном для синтеза цепи А.
Полуконсервативный синтез. Если разделить цепи А и А' одной молекулы ДНК, то каждая из них будет служить матрицей для синтеза соответствующей недостающей цепи. Новые молекулы ДНК будут содержать одну новую и одну исходную материнскую цепь ДНК. Две дочерние молекулы ДНК полностью идентичны исходной материнской.
Антипараллельность. Две цепи в молекуле ДНК антипараллельны. Это значит, что у нуклеотида в начале одной цепи находится остаток дезоксирибозы со свободной гидроксогруппой (-OH) у 3'-атома углерода, а у комплементарного ему нуклеотида в начале другой цепи находится остаток фосфорной кислоты, соединенный с 5'-атомом углерода дезоксирибозы. Соответственно, первая цепь заканчивается нуклеотидом с 5'-концом, а вторая цепь — нуклеотидом с 3'-концом.
Челночный синтез. Фермент ДНК-полимераза, обеспечивающий синтез новой цепи ДНК на матрице исходной цепи, соединяет нуклеотиды только в направлении от 5'-нуклеотида к 3'-нуклеотиду. Поэтому синтез идет на одной цепи вперед в направлении 5'—3' непрерывно, а затем по другой цепи назад к 5'-концу фрагментами. Отдельные фрагменты соединяются позже.
Процесс репликации протекает в три стадии.
Стадия I — инициация. Репликация молекулы ДНК начинается с разъединения двойной спирали с одного конца, причем процесс идет не на всем участке молекулы, а частями, фрагментарно. Эта реакция проводится в присутствии нескольких белковых факторов. Две цепи в молекуле ДНК связаны достаточно прочно, поэтому для их разъединения необходимы специальные белки. Белки первого типа, перемещаясь по молекуле ДНК, раскручивают спираль и разрушают водородные связи между комплементарными основаниями, расплетая двойную спираль. Белки второго типа предотвращают повторное соединение двух цепей и обеспечивают эффективность действия белков первого типа. Кроме того, они выпрямляют одиночные цепи ДНК и обеспечивают продвижение фермента ДНК-полимеразы, который катализирует синтез новых цепей. В результате действия всех белковых факторов образуется репликационная вилка (рис. 25).
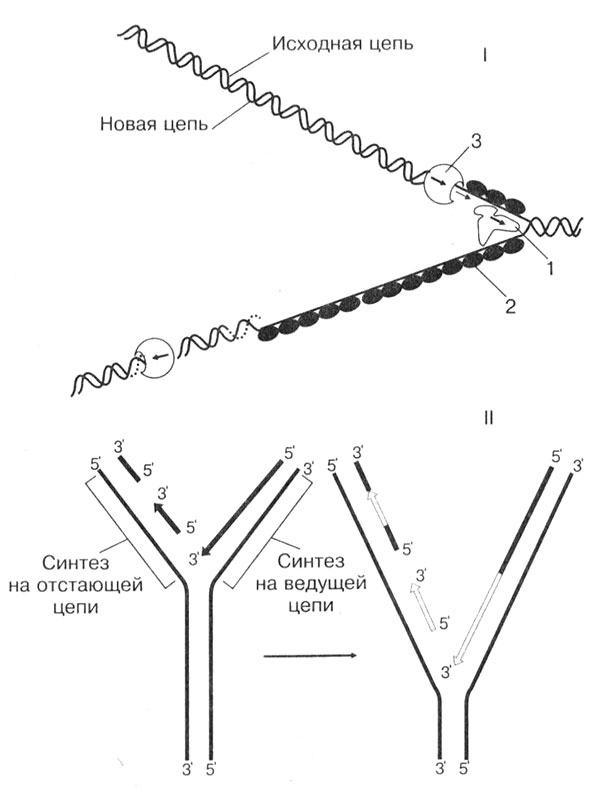
Рис. 25. Репликация ДНК: I — строение репликационной вилки: 1 — ДНК-расплетающий белок; 2 — ДНК-связывающий белок; 3 — ДНК-полимераза; II — синтез ДНК на ведущей и отстающей цепи (стрелками показано направление синтеза и перемещение ДНК-полимеразы)
Стадия II — элонгация. На этой стадии происходит собственно синтез ДНК. По принципу комплементарности на каждой из цепей ДНК выстраиваются нуклеотиды. Самокопирующий фермент ДНК-полимераза, передвигаясь по репликационной вилке, соединяет между собой нуклеотиды в направлении от 5'- к 3'-концу. Движение фермента идет только в одном направлении. Так как цепи ДНК антипараллельны, то по другой цепи ДНК-полимераза движется в обратном направлении и синтезирует фрагмент от начала репликационной вилки к концу цепи.
На первой цепи фермент, дойдя до начала репликационной вилки, как бы стимулирует движение связывающего белка. Он передвигается выше, расплетая молекулу ДНК дальше. Фермент вновь передвигается по первой цепи, продолжая прерванный синтез с последнего нуклеотида. На другой же цепи фермент синтезирует второй фрагмент ДНК в обратном направлении.
Репликационная вилка оказывается несимметричной. Одна из дочерних цепей ДНК (ведущая) строится непрерывно, а другая (отстающая) — синтезируется прерывисто, в виде отдельных фрагментов. Фрагменты соединяются друг с другом позже, только после синтеза следующих фрагментов.
Последняя III стадия называется терминацией. С вновь синтезированных молекул ДНК снимаются все белковые факторы, ферменты. Две дочерние молекулы ДНК расходятся, спирализуются и приобретают соответствующую структуру. Процесс синтеза ДНК заканчивается.
Копирование ДНК происходит с высокой точностью. В среднем на каждые 1 ? 109 комплементарных пар нуклеотидов, образующихся в ходе репликации, приходится одна ошибка. Эти ошибки устраняются особой корректирующей системой белков, распознающих и удаляющих неправильные нуклеотидные остатки.
Точность копирования обеспечивает правильность передачи наследственной информации. Весь процесс репликации происходит за счет энергии АТФ.






