Жгутики
По механизму движения бактерии подразделяют на плавающие и скользящие, или ползающие. Последние активно передвигаются по плотной поверхности благодаря волнообразным сокращениям тела (некоторые виды Mycoplasma, Myxococcus и др.). У плавающих бактерий органом движения являются жгутики, которые представляют собой тонкие длинные нитевидные белковые образования диаметром 12 – 30 нм и длиной от 6 – 9 до 80 мкм. Белок, из которого построены жгутики, получил название флагеллина. Он отличается от других белков, содержащихся в бактериальной клетке. Флагеллин обладает сократительной способностью, хотя механизм ее не совсем понятен.
Жгутик состоит из однотипных спиралевидно или продольно уложенных вокруг полой сердцевины белковых субъединиц, образующих цилиндрическую структуру, которая особым образом прикреплена к бактериальной клетке. По характеру расположения жгутиков и их количеству подвижные бактерии условно делят на четыре группы (рис. 11):
1) монотрихи – один полярно расположенный жгутик (Vibrio cholerae);
2) лофотрихи – пучок жгутиков на одном конце (Pseudomonas methanica);
3) амфитрихи – пучки жгутиков на обоих концах клетки (Spirillum volutans);
4) перитрихи – множество жгутиков, расположенных вокруг клетки (E. coli, Salmonella typhi).
Жгутик состоит из трех компонентов – спиральной жгутиковой нити постоянной толщины, крючка и базального тельца (рис. 12). Крючок, к которому присоединена жгутиковая нить, имеет длину 30 – 45 нм и состоит из отличающегося от флагеллина белка. Он соединен с базальным тельцем, которое располагается целиком в оболочке (в клеточной стенке и ЦМ).
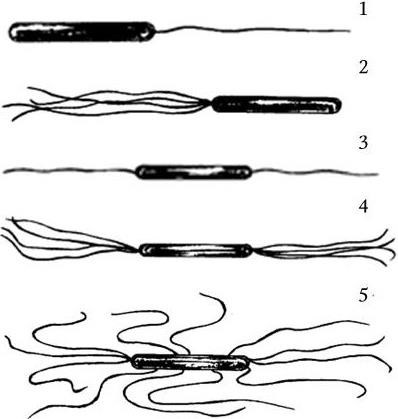
Рис. 11
1 – монотрихи; 2 – лофотрихи; 3, 4 – амфитрихи; 5 – перитрихи
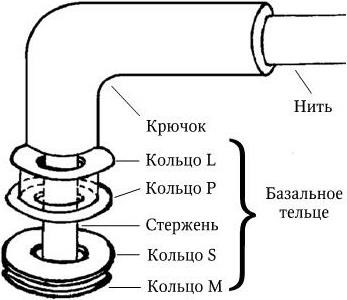
Рис. 12
Базальное тельце состоит из центрального стержня, заключенного в систему особых колец. У грамотрицательных бактерий их две пары: внешняя (кольца L и P) и внутренняя (кольца S и М). Кольца L и P расположены внутри клеточной стенки (кольцо L – в ЛПС, а кольцо P – в слое пептидогликана). Они выполняют, очевидно, роль втулки для стержня. Внутренняя пара (кольца S и M) фиксирована на ЦМ, причем кольцо S располагается в периплазматическом пространстве, а кольцо М – на ЦМ или в ней.
Жгутики у грамположительных бактерий, имеющих более толстую и гомогенную клеточную стенку, содержат только одну пару колец – S и M. Вращение жгутиков в клеточной стенке происходит из-за вращательного движения колец S и M относительно друг друга и обеспечивается за счет энергии трансмембранного градиента ионов водорода или натрия. Благодаря такому вращению происходит движение бактерий в наиболее благоприятном для них направлении. Жгутиковый аппарат обладает особым бинарным переключателем, который позволяет менять направление вращения жгутиков против часовой стрелки на противоположное. Таким способом бактерии, получив химический сигнал из окружающей среды, изменяют направление движения и выбирают оптимальные условия обитания. По всей вероятности, базальное тельце (его внутреннее кольцо М) непосредственно связано с какими-то дополнительными жгутиковыми белками, которые необходимы для сборки жгутиков и управления переключением направления их вращения и которые расположены либо в ЦМ, либо сразу под ней. Со жгутиковым аппаратом связана также и хемотаксическая активность таких бактерий. Генетический контроль синтеза жгутиковых белков, их сборки и активности осуществляется особым опероном. Установлено, что мутации в области mot-генов (англ. motility – подвижность) приводят к потере только подвижности, однако все структуры жгутиков сохраняются; мутации в cheгенах (англ. chemotaxis – хемо + подвижность) – к потере хемотаксической активности при сохранении структуры жгутиков и их подвижности. Подвижность бактерий определяют либо микроскопически (с помощью фазово-контрастной или обычной световой микроскопии «раздавленной» или «висячей» капли соответственно), либо бактериологически (при посеве уколом в столбик полужидкого агара: подвижные бактерии дают диффузный рост, а неподвижные – растут только по ходу укола). Жгутики хорошо выявляются при электронной микроскопии (рис. 13). Жгутиковые бактерии могут двигаться с большой скоростью, например Bacillus megaterium движется со скоростью 27 мкм/с, а Vibrio cholerae – 200 мкм/с.
Донорные ворсинки. У бактерий, являющихся носителями конъюгативных плазмид (F-плазмид, R-плазмид и др.), имеются длинные (0,5 – 10 мкм) нитевидные структуры белковой природы, получившие название донорных ворсинок, или донорных пилей (англ. pile – волосок). Как и жгутики, они имеют внутреннюю полость и построены из особого белка. Их синтез находится под контролем плазмидных генов. Они служат аппаратом конъюгации – с их помощью устанавливается непосредственный контакт между донорной и реципиентной клетками. Донорные пили обнаруживают с помощью донорспецифических фагов, которые на них адсорбируются и далее вызывают лизис клетки-хозяина. Донорные пили встречаются в количестве 1 – 2 на клетку.
Фимбрии, или реснички. Фимбрии (англ. fimbria – бахрома) – короткие нити, в большом количестве (до многих тысяч) окружающие бактериальную клетку (рис. 14). Подобно жгутикам и донорным ворсинкам, они прикреплены к клеточной стенке, но значительно короче и тоньше – их длина 0,1 – 12,0 мкм, диаметр 25 нм. Белок фимбрий отличается от белков жгутиков и донорных ворсинок. Биологическое значение фимбрий, по-видимому, состоит в том, что с их помощью бактерии прикрепляются к определенным поверхностям. Для многих патогенных бактерий фимбрии являются важными факторами патогенности, так как с их помощью бактерии прикрепляются к чувствительным клеткам и заселяют их, т. е. фимбрии служат для бактерий факторами адгезии и колонизации.
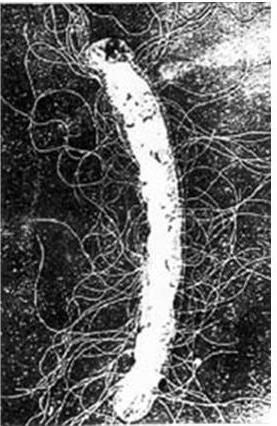
Рис. 13

Рис. 14






