Вирусный гепатит В
Гепатит В – инфекционное заболевание человека, характеризующееся избирательным поражением печени вирусом. Эта форма гепатита является наиболее опасной по своим последствиям среди всех известных форм вирусных гепатитов. Его возбудителем является вирус гепатита В (HBV).
Впервые антиген вируса гепатита В был обнаружен Б. Блюмбергом в 1964 г. в сыворотке крови австралийского аборигена, а сам возбудитель был обнаружен в 1970 г. Д. Дейном [и др.] и получил название частиц Дейна, поскольку не было полной уверенности в том, что это действительно вирус, а не его компоненты. В последующем все сомнения отпали, так как в составе частиц Дейна были обнаружены геномная ДНК и вирусная ДНК-зависимая ДНК-полимераза. В составе вириона имеются три основных антигена, для которых в 1974 г. были введены следующие обозначения:
1. HBsAg – поверхностный (superficial), или растворимый (soluble), или австралийский антиген.
2. HBcAg – сердцевинный антиген (сor-антиген).
3. HBeAg – антиген e, локализован в сердцевине вириона и, в отличие от HBcAg, не только присутствует в составе вириона, но и циркулирует в крови в свободном виде или в виде комплекса с антителом анти-HBeAg. Он выделяется в кровь из гепатоцитов при активной репликации HBV.
Поверхностный антиген – HBsAg – существует в виде трех морфологически различных вариантов: 1) представляет суперкапсид цельного вириона; 2) в большом количестве встречается в виде частиц диаметром 20 нм, имеющих сферическую форму; 3) в виде нитей длиной 230 нм. Химически они идентичны. В составе HBsAg имеется один общий антиген а и две пары взаимоисключающих типоспецифических детерминантов: d/y и w/r, поэтому существуют четыре основных субтипа HBsAg (и соответственно HBV): adw, adr, ayw и ayr. Антиген а обеспечивает формирование общего перекрестного иммунитета ко всем субтипам вируса.
Собственно вирион – частица Дейна – имеет сферическую форму и диаметр 42 нм. Суперкапсид вириона состоит из трех белков: главного (основного), большого и среднего (рис. 88, 1). Геном заключен в капсид и представлен двунитевой кольцевидной ДНК с м. м. 1,6 МД. ДНК состоит приблизительно из 3200 нуклеотидов, однако ее «плюс» – нить на 20 – 50 % короче «минус» – нити. С 5'-концом длинной нити ковалентно связан вирусспецифический белок. 5'-концы обеих нитей комплементарны и образуют «липкие» последовательности длиной в 300 нуклеотидов, благодаря чему нити замыкаются в кольцо. Содержание Г + Ц в вирионной ДНК 48 – 49 мол %. В сердцевине вириона находится кроме геномной ДНК-вирусная ДНК-зависимая ДНК-полимераза. «Минус» – нить ДНК HBV содержит всего четыре гена (S, C, P и X), но они организованы очень компактно (рис. 88, 2). Гены S, C, P, X сильно перекрываются и контролируют синтез следующих продуктов. Ген S кодирует синтез главного белка оболочки и содержит всю информацию о поверхностном антигене HBsAg. Кроме того, он кодирует синтез среднего и большого белков оболочки. Белки содержат общий СООН-конец, но их трансляция начинается с трех различных инициаторных кодонов. Ген С кодирует синтез капсидных белков (HBcAg и HBeAg); хотя эти белки кодируются одним геном, пути их трансляции различны. Ген Р – самый большой. Он включает в себя часть всех трех других генов и кодирует ферменты, необходимые для репликации вируса. В частности, он кодирует обратную транскриптазу, домен фермента РНК-азы Н, 5'-концевой белок «минус» – цепи. Ген X кодирует белки, регулирующие экспрессию (выражение) всех вирусных генов, в частности белок с м. м. 17 кД, который является трансактиватором транскрипции генов.
Белки, образующие поверхностный антиген, существуют в гликозилированной (gp) и негликозилированной форме. Гликозилированными являются gp27, gp33, gp36 и gp42 (цифры обозначают м. м. в кД). Суперкапсид HBV состоит из главного, или основного, S-белка (92 %); среднего М-белка (4 %) и большого, или длинного, L-белка (1 %).
Главный белок – p24/gp27, или основной белок (белок S), является основным компонентом оболочки HBV. В отсутствие других оболочечных белков он полимеризуется и образует сферические частицы диаметром 20 нм, которые состоят из 100 полипептидных молекул.
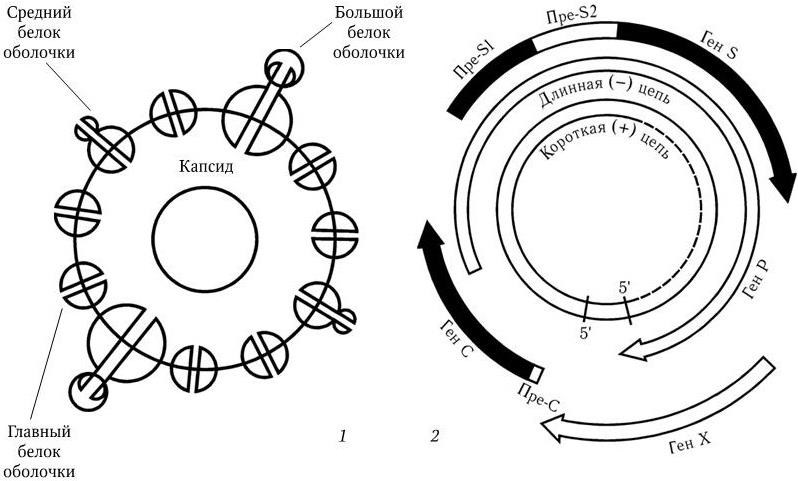
Рис. 88
Большой белок – p39/gp42, или длинный белок (белок L), присутствует во всех трех формах HBsAg. Он играет важную роль в морфогенезе вирионов и в выходе их из клетки. L-белок содержит последовательность белка М, которая на N-конце дополнена последовательностями из 108 (ayw) или 119 (adw, adr, ayr) аминокислотных остатков, кодируемых пре-S1-областью S-гена.
Средний белок – gp33/gp36, или белок М, также присутствует во всех трех морфологических формах HBsAg. Белок М содержит на N-конце участок из 55 аминокислотных остатков, кодируемых пре-S2-областью S-гена. Предполагается, что этот участок играет важную роль в распознавании вирусом гепатита В клеток печени ограниченного круга хозяев (человек, обезьяна шимпанзе). Последовательности белков, кодируемых пре-S-областями S-гена, обладают высокой иммуногенностью, а их детерминанты расположены на поверхности вириона. Поэтому антитела против этих антигенов играют важную роль в формировании иммунитета против гепатита В.
Синтез вирусных белков жестко контролируется на уровне транскрипции и трансляции. При транскрипции вирусного генома синтезируются два типа мРНК: а) меньшая – 2100 нуклеотидов – кодирует главный и средний белки оболочки;
б) бо?льшая – 3500 нуклеотидов, т. е. длиннее само?й геномной ДНК; она содержит концевые повторы длиной 100 нуклеотидов. Этот вид мРНК кодирует белок капсида и продукты гена Р. Она также является матрицей для репликации вирусной ДНК. В составе генома есть энхансеры (усилители транскрипции) – регуляторные элементы, которые активируют экспрессию всех вирусных генов и действуют преимущественно в клетках печени. В частности, ген S экспрессируется на очень высоком уровне только в клетках печени и под влиянием стероидных гормонов. Это обстоятельство и объясняет, почему хронический гепатит В и рак печени (гепатома) наблюдаются у мужчин чаще, чем у женщин, у которых уровень стероидных гормонов ниже.
Другие регуляторные элементы вируса гепатита В модулируют (контролируют) уровни синтеза отдельных белков. Например, большой белок синтезируется лишь в малом количестве. Больше всего его на поверхности инфекционных вирионов. А главный белок и, в меньшей степени, средний белок синтезируются в огромном количестве и покидают клетки в составе частиц поверхностного антигена, которых в сыворотке крови содержится во много раз больше, чем зрелых вирионов. Количество частиц поверхностного антигена может составлять 1011– 1013 на 1 мл крови (несколько сотен мкг).
Вирус гепатита В выделен в новое семейство вирусов – Hepadnaviridae, род Orthohepadnavirus. Сходные с ним гепаднавирусы обнаружены у различных животных (земляных белок, сурков, бурундуков, пекинских уток).
Репродукция гепаднавирусов происходит несколько необычным образом. В частности, репликация геномной ДНК происходит через промежуточное звено – РНК, т. е. с механизмом обратной транскрипции.
Жизненный цикл вируса гепатита В.
1. Адсорбция на клетке.
2. Проникновение в клетку с помощью механизма рецепторопосредованного эндоцитоза (окаймленная ямка ? окаймленный пузырек ? лизосома ? выход нуклеокапсида и проникновение вирусного генома в ядро гепатоцита).
3. Внутриклеточное размножение.
В ходе проникновения в клетку происходит удлинение (достраивание) короткой («плюс») цепи ДНК. В ядре клеточная ДНК-зависимая РНК-полимераза синтезирует РНК размером 3500 нуклеотидов (прегеном) и мРНК, меньшие по размерам, для синтеза вирусных белков. Затем прегеном и вирусная ДНК-полимераза упаковываются во вновь синтезированный капсид, который переносится в цитоплазму. Здесь и происходит обратная транскрипция прегенома. На нем синтезируется новая «минус» – нить ДНК. После завершения синтеза «минус» – нити ДНК прегеномная РНК разрушается. Вирионная ДНК-полимераза на «минус» – цепи синтезирует «плюс» – цепь. Вирусная ДНК, теперь уже двухцепочечная, может существовать в клетке довольно долго и возвращаться в ядро для следующего цикла репликации. Если новая вирусная частица не подвергается дальнейшей репликации, то сформировавшийся нуклеокапсид, проходя через мембрану клетки, покрывается суперкапсидом, отпочковывается от клетки, и в нем немедленно прекращается удлинение короткой «плюс» – цепи ДНК. Вот почему длина этой нити варьирует. При типичной острой форме гепатита В в крови последовательно появляются следующие серологические маркеры: HBsAg, HBeAg и антитела (IgM, IgG): анти-HBcAg, анти-HBeAg и анти-HBsAg.
В составе вируса гепатита В нет онкогена, однако установлено, что, внедряясь в клеточную хромосому (в разные ее участки), вирусная ДНК может индуцировать в них различные генетические перестройки – делеции, транслокации, амплификации, которые и могут стать причиной развития рака печени – одного из самых тяжких последствий вирусного гепатита В.
Резистентность. Вирус гепатита В обладает высокой устойчивостью. При комнатной температуре сохраняет жизнеспособность в течение 3 мес., в замороженном состоянии – несколько лет. Вирус полностью инактивируется при автоклавировании (120 °C), при кипячении в течение 30 мин, сухим жаром при температуре 180 °C в течение 60 мин, при 60 °C – в течение 10 ч. Устойчив в кислой среде, но разрушается в щелочной. Вирус погибает при обработке Н2О2, хлорамином, формалином, фенолом и при УФ-облучении.
Эпидемиология. Источником заражения вирусом гепатита В является только человек. Вопреки прошлым представлениям о том, что заражение вирусом гепатита В
происходит исключительно парентеральным путем, теперь доказано, что он обнаруживается в различных секретах и экскретах: в слюне, носоглоточных выделениях, испражнениях, слезной жидкости, в сперме, менструальной крови и пр. Таким образом, заражение происходит не только парентеральным путем, но и половым, и вертикальным (от матери плоду), т. е. практически заражение вирусом гепатита В возможно разными способами.
От гепатита B в мире погибло столько же людей, сколько за все годы Второй мировой войны. Число носителей HBV, по данным ВОЗ, составляет от 0,1 до 20 % населения разных стран или регионов.
Патогенез и клиника. Вирус гематогенным путем заносится непосредственно в печень. В патогенезе гепатита важную роль играют аутоиммунные гуморальные и клеточные реакции. Предполагается, что поражение гепатоцитов связано не столько с непосредственным действием самого вируса, сколько с иммунологическими реакциями хозяина, возникающими в связи с модификацией клеточной мембраны вирусными белками, которые индуцируют появление аутоантител к клеткам печени. Поэтому развивающийся хронический гепатит и цирроз печени можно рассматривать как аутоиммунное заболевание.
Клеточные аутоиммунные реакции к вирусным белкам, содержащимся в мембране гепатоцитов, опосредуются Т-цитотоксическими лимфоцитами и другими киллерными клетками печени. Поэтому острую дистрофию печени можно рассматривать как реакцию отторжения своеобразного гетеротрансплантата.
Инкубационный период длится от 45 до 180 дней, в среднем составляет 60 – 90 дней. Клиническое течение гепатита В характеризуется большим разнообразием;
болезнь может протекать: в латентной форме, выявляемой лишь лабораторными методами, в типичной желтушной форме и в злокачественной форме, заканчивающейся летально. Продолжительность преджелтушной стадии составляет от одного дня до нескольких недель. Желтушный период, как правило, длительный и характеризуется хорошо выраженными симптомами (желтуха, гипербилирубинемия, потемнение мочи, желтушность склер). Затяжная форма наблюдается у 15 – 20 % больных, а у 90 % из них развивается хронический гепатит В. У больных с затяжной формой нередко наблюдаются аутоиммунные процессы, сопровождающиеся повышенным содержанием противопеченочных антител, которые выявляются с помощью иммуносорбентного анализа (ИФМ). У детей гепатит В протекает в более легкой форме и часто без развития желтухи, у детей младшего возраста – преимущественно бессимптомно.
Постинфекционный иммунитет (гуморальный и клеточный) длительный, пожизненный, обусловлен вируснейтрализующими антителами (анти-HBsAg) при отсутствии в крови поверхностного антигена. Нередко наблюдается скрытая иммунизация вследствие повторного контакта с HBV, которая является причиной широкого распространения иммунитета к вирусу среди населения. Обычно больные с острой формой гепатита В выздоравливают полностью по мере накопления антител к нему. Однако в некоторых случаях, несмотря на высокий уровень вирусного антигена в крови (обстоятельство, объясняющее, почему парентеральное заражение происходит наиболее часто), антитела к нему не вырабатываются. Вирус сохраняется в печени, и человек на долгое время, иногда пожизненно, становится хроническим носителем. Это обстоятельство связано, очевидно, со слабым иммунным ответом. Одним из наиболее частых исходов хронического гепатита В является цирроз печени и рак печени, который развивается по истечении латентного периода продолжительностью до 30 – 50 лет.
Методы диагностики гепатита В. В настоящее время основным методом диагностики гепатита В является использование реакции обратной пассивной гемагглютинации (РОПГА) для обнаружения вируса или его поверхностного антигена – HBsAg. Как уже отмечалось, в крови поверхностного антигена содержится во много раз больше, чем самого вируса (в 100 – 1000 раз). Для реакции РОПГА используют сенсибилизированные антителами против вируса гепатита В эритроциты. При наличии антигена в крови происходит реакция гемагглютинации. РОПГА проста, удобна, очень специфична. Для обнаружения антител к вирусному антигену HBsAg используют различные иммунологические методы (РСК, РПГА, ИФМ, РИМ и др.). Кроме того, для обнаружения HBV и его антигенов используют варианты ПЦР.
Для обнаружения в сыворотке больного антител к вирусному антигену (HBsAg) могут быть использованы различные иммунологические методы (РСК, РПГА, реакция преципитации, ИФМ, РИМ и др.).
Специфическая профилактика. Принимая во внимание высокий уровень заболеваемости гепатитом В, а также, что в мире очень много носителей HBV, по рекомендации ВОЗ прививки против гепатита В являются обязательными и должны проводиться на первом году жизни. Для вакцинации предложено два типа вакцин. Для приготовления одной из них в качестве сырья используют плазму вирусоносителей, поскольку в ней вирусный антиген содержится в количествах, достаточных для приготовления вакцины. Главное условие для приготовления этого типа вакцин – их полная безопасность, т. е. полная инактивация вируса, что и предусматривается технологией приготовления вакцины. Для изготовления вакцины другого типа применяют методы генной инженерии, в частности, для получения антигенного материала используют рекомбинантный клон дрожжей, вырабатывающих поверхностный антиген вируса гепатита В. О дивакцине против гепатитов A и B см. с. 6.
Обе вакцины обладают высокой эффективностью (защищают 95 % привитых). Продолжительность поствакцинального иммунитета не менее 5 – 6 лет. В России созданы вакцины как для взрослых людей, так и для новорожденных и детей раннего возраста – наиболее важный компонент борьбы с гепатитом В в глобальном плане. Полный курс прививки состоит из трех инъекций:
I доза – сразу после рождения; II доза – через 1 – 2 мес.; III доза – до конца 1-го года жизни.
Эти прививки включены в расширенную программу иммунизации ВОЗ и совмещаются с календарем ее проведения (по рекомендации ВОЗ, на 1-м году жизни проводят прививки против туберкулеза, полиомиелита, гепатита В, кори, столбняка, дифтерии, коклюша).
Гамма-глобулин, содержащий антитела против HBV, применяют для экстренной пассивной иммунопрофилактики лицам, имевшим контакт с больным гепатитом В. Для лечения гепатита B (острой и хронической форм) используют интерферон и амиксин (для индукции его эндогенного синтеза). При лечении хронического гепатита B эффективен новый препарат ламивудин (синтетический нуклеозид).






