Репликация вирусных геномов
Тип геномной вирусной ДНК определяет особенности ее репликации.
1. Двунитевая ДНК – репликация происходит по обычному механизму полуконсервативной репликации: нити разделяются, и на каждой из них достраивается комплементарная ей нить.
2. Однонитевая ДНК. Ее репликация происходит через образование вначале репликативной формы, а затем промежуточной репликативной формы. Репликативная форма возникает в результате синтеза на исходной вирионной ДНК («+» нити) комплементарной ей « – » нити, т. е. однонитевая ДНК превращается в двунитевую структуру ДНК. Промежуточная репликативная форма – это репликативная форма, « – » нить которой служит матрицей для синтеза «+» нити ДНК, идентичной исходной вирионной ДНК. Такой механизм обеспечивает передачу генов дочерним вирионам (рис. 80. I).
3. У вирусов, геном которых представлен однонитевой РНК, ее репликация происходит по следующей схеме: вначале на вирионной РНК (вРНК) синтезируются комплементарные ей РНК (кРНК). Этот процесс катализируется специфической РНК-репликазой I. Затем на кРНК синтезируется комплементарная ей, но идентичная исходной вирионная РНК (вРНК), этот процесс также катализируется специфической репликазой II. Таким образом, репликация идет по схеме (рис. 80. II):
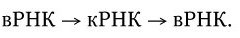
4. Репликация однонитевой РНК ретровирусов происходит с участием обратной транскриптазы. Вначале на вРНК обратная транскриптаза синтезирует комплементарную ей «минус» цепь ДНК, а затем на ней – «плюс» нить ДНК. Двунитевая ДНК интегрируется в хромосому клетки и там служит матрицей для синтеза разных классов вирусных РНК. Таким образом, репликация ретровирусов происходит по схеме:
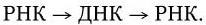
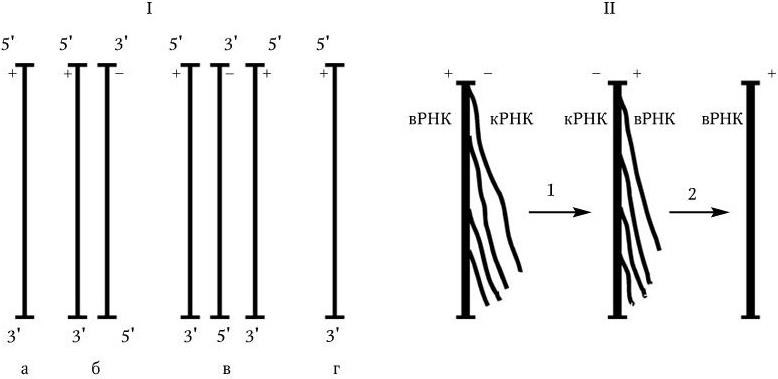
Рис. 80
а – геномная однонитевая ДНК; б – репликативная форма; в – промежуточная репликативная форма; г – однонитевая дочерняя геномная ДНК; вРНК – вирионная (геномная) РНК; кРНК – комплементарная РНК. Объяснение в тексте
5. Размножение вируса гепатита В также протекает с участием обратной транскриптазы, но вначале клеточная РНК-полимераза синтезирует на вирусной ДНК прегеномную РНК, после чего вирусная ревертаза синтезирует на ней минус-цепь ДНК, которая достраивается плюс-цепью ДНК. В виде двунитевой ДНК вирус интегрируется в хромосому клетки-хозяина, где на ней транскрибируется вирусная РНК.
Существуют некоторые общие закономерности размножения вирусов. Во-первых, все РНК-содержащие вирусы, кроме вирусов гриппа и ретровирусов, размножаются в цитоплазме. Для своего размножения вирусы гриппа А и В и ретровирусы проникают в ядро, что связано с особенностями поведения их генома. Во-вторых, размножение всех ДНК-содержащих вирусов, кроме вирусов оспы, протекает в ядре, где происходит транскрипция и репликация их геномных нуклеиновых кислот, и в цитоплазме, где происходит трансляция вирусных белков, их процессинг и морфогенез вирионов. Лишь размножение вирусов группы оспы происходит в цитоплазме клетки, поскольку они обладают собственными системами транскрипции.
Другая особенность размножения вирусов заключается в том, что их нуклеокапсидные белки синтезируются на свободных полирибосомах (не связанных с мембраной), а суперкапсидные белки – на рибосомах, ассоциированных с мембранами (на шероховатых мембранах). Кроме того, белки некоторых вирусов подвергаются протеолитическому процессингу и гликозилированию. Различают два типа протеолитического процессинга: каскадный и точечный.
При каскадном протеолизе вновь синтезированный вирусный полипептид-предшественник (полипротеин) подвергается последовательному «нарезанию» с образованием более коротких полипептидов, часть из которых дополнительно разрезается на более мелкие субъединицы. Ряд ступеней такого каскадного протеолиза осуществляется определенной областью самого полипротеина, обладающего протеазной активностью. Такому каскадному процессингу подвергаются белки у ретровирусов, пикорнавирусов, aльфа-, флави– и других вирусов. Для них такое протеолитическое нарезание белков является жизненно важным этапом репродукции, поскольку оно обусловливает реализацию их функций.
При точечном протеолизе разрезанию подвергается один (реже несколько) из вирусных полипептидов. Разрезание происходит, как правило, в определенном участке полипептида. Такой тип протеолиза необходим для того, чтобы определенный белок вируса приобрел свою специфическую активность. Например, суперкапсидный белок вируса гриппа – гемагглютинин – разрезается на две субъединицы: боEльшую и меньшую. В результате меньшая субъединица приобретает способность сливаться с мембранами клетки-мишени и ее лизосомами. Благодаря этому вирус гриппа приобретает способность проникать в клетку. Такой точечный протеолиз наблюдается у ортомиксовирусов, парамиксовирусов, ротавирусов, вирусов группы оспы и др. Точечный протеолиз, как и каскадный, жизненно важен для вируса.
Наконец, еще одна особенность вирусов, обладающих суперкапсидом, заключается в том, что суперкапсидные белки подвергаются в ходе своей транспортировки на наружную поверхность клеточной мембраны гликозилированию.






