Механизмы питания бактерий
Большинство бактерий живет в среде, мало подходящей для того, чтобы поддерживать строгое соотношение воды, солей и органических веществ, без которого невозможна жизнь. Это обусловливает необходимость постоянного и тщательного регулирования обмена различными веществами, который происходит между клеткой и внешней средой и контролируется клеточной мембраной. Она проницаема для многих веществ, поток их идет в обоих направлениях (из клетки и в клетку), но структура мембраны такова, что она обладает избирательной и неравномерной проницаемостью, определяющей механизмы питания бактерий.
Питательные вещества из внешней среды поступают в бактериальную клетку с помощью трех основных механизмов: пассивной диффузии, облегченной диффузии и активного транспорта (рис. 17).
Пассивная диффузия осуществляется за счет различного содержания питательных веществ в среде и в клетке и происходит в направлении от большей концентрации к меньшей, т. е. по градиенту концентрации. Когда концентрация вещества по ту и другую сторону мембраны уравнивается, пассивная диффузия прекращается. Ее скорость зависит от величины градиента, но она имеет определенный предел. Таким путем в клетку проникает (и покидает ее) вода вместе с растворенными в ней различными мелкими молекулами, способными проходить через мелкие поры мембраны. Для пассивной диффузии характерно отсутствие субстратной специфичности, и она не требует затраты энергии.
Облегченная диффузия характеризуется выраженной субстратной специфичностью и протекает при обязательном участии специфических белков, локализованных в мембране; синтез некоторых из них индуцируется соответствующими субстратами. Эти белки, получившие название пермеаз (англ. permeate – проникать, проходить сквозь), обладают субстратной специфичностью. Они распознают и связывают молекулу субстрата на внешней стороне мембраны и обеспечивают каким-то образом ее перенос через мембрану. На внутренней поверхности мембраны комплекс пермеаза – субстрат диссоциирует, освободившаяся молекула субстрата включается в общий метаболизм клетки, а пермеаза повторяет очередной цикл переноса своего субстрата, который не способен проникать через мембрану путем простой диффузии. Главное свойство пермеаз – способность проходить через мембрану как с присоединенной молекулой субстрата, так и без нее. Однако облегченная диффузия происходит только по градиенту концентрации, но не против него, поэтому она не требует затраты энергии. Пермеазы, присоединившись к субстрату, повышают его способность проникать через мембрану. Облегченная диффузия протекает со значительно более высокой скоростью, чем пассивная. Ее скорость подчиняется закону Михаэлиса – Ментен, и при достижении равновесия концентрация субстрата, доставляемого посредством облегченной диффузии, на внутренней и внешней поверхностях мембраны становится одинаковой.
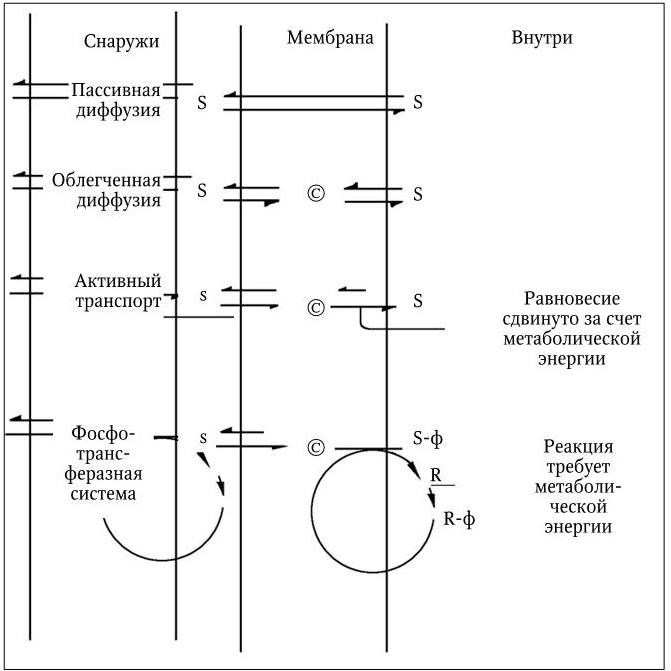
Рис. 17
Разная длина стрелок указывает на сдвиг равновесия реакции в сторону более длинной стрелки. S и s означают соответственно высокую и низкую концентрации растворенных веществ; © – белок-переносчик (пермеаза); R – белок HРr; R-ф – фосфо-HРr; ф – фосфатная группа
Активный транспорт. С помощью механизмов активного транспорта растворенные вещества могут поступать в клетку против градиента концентрации, поэтому активный транспорт требует от клетки затраты энергии. У бактерий этот механизм питания является преобладающим. С его помощью они обеспечивают такие концентрации растворенных питательных веществ внутри клетки, которые могут во много раз превышать их концентрации во внешней среде и обеспечивают им высокие скорости метаболизма даже при низкой концентрации химических веществ в окружающей среде. У многих бактерий, в особенности грамотрицательных, в активном транспорте принимают участие особые связывающие белки, не идентичные пермеазам и не входящие в структуру мембраны, а локализованные в периплазматическом пространстве. У связывающих белков отсутствует каталитическая активность, но они обладают очень высоким сродством к определенным питательным веществам – к различным аминокислотам, сахарам, неорганическим ионам. Выделено и изучено более 100 различных связывающих белков, которые образуют прочные комплексы со своими субстратами и необходимы для их активного переноса через мембрану. Связывающие белки функционируют только в комплексе со специфическими пермеазами, осуществляющими активный перенос субстрата через мембрану. Метаболическая энергия, необходимая для этого, используется для снижения сродства пермеазы к своему субстрату на внутренней поверхности мембраны по сравнению с ее сродством к нему на внешней стороне мембраны. В результате этих превращений происходит изменение скорости выхода субстрата наружу, она становится во много раз меньше скорости его поступления в клетку. При этом механизме активного транспорта через мембрану в клетку поступают против градиента концентрации химически не измененные питательные вещества. У бактерий, вместе с тем, существуют и такие транспортные системы, которые переводят питательные вещества в химически измененную форму, не способную проникать через мембрану. К их числу относится фосфотрансферазная система, широко распространенная среди бактерий. С помощью этой системы транспортируются многие сахара и их производные, в процессе переноса они фосфорилируются и поступают в клетку в виде сахарофосфатов. Поскольку мембрана для последних непроницаема, сахарофосфаты остаются внутри клетки.
Фосфотрансферазная система состоит из двух неспецифических компонентов: ферментов I и HPr и набора субстрат-специфических белков, связанных с мембраной и обозначенных как ферменты II. Фермент I обеспечивает перенос богатой энергией фосфатной группы от фосфоенолпирувата на гистидиновый остаток фермента HPr, который превращается в фосфо-HPr. Последний является общим донором фосфорильной группы для всех субстратов, переносимых фосфотрансферазной системой. Фосфорилирование же их осуществляется субстрат-специфическими белками из группы ферментов II, которые выполняют также и функции пермеаз. У мутантных бактерий, лишенных фермента I или белка HPr, ферменты II осуществляют облегченную диффузию своих субстратов.
Транспортные системы в жизни клетки выполняют две основные функции:
1) поддерживают на высоком уровне внутриклеточные концентрации всех субстратов, необходимых для осуществления важнейших биохимических реакций с максимальными скоростями даже при низких концентрациях этих химических веществ во внешней среде;
2) регулируют внутриклеточное осмотическое давление, поддерживают оптимальную для метаболической активности концентрацию растворенных веществ (небольших молекул и ионов).