Рост и размножение бактерий
Различают рост индивидуальных клеток и рост популяции. Каждый из них характеризуется своими особенностями и закономерностями. Под ростом индивидуальной клетки понимают увеличение ее биомассы, наступающее в результате синтеза клеточного материала. Объем клетки можно вычислить, если известны ее продольные и поперечные размеры. Для шаровидных клеток он определяется по формуле:

а для цилиндрических
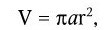
где r – радиус клетки; a – длина клетки.
Рост палочек происходит в длину, поэтому удельная поверхность (отношение поверхности к объему) остается примерно постоянной, и скорость роста в определенных условиях может быть постоянной.
У сферических клеток рост происходит во всех направлениях, поэтому удельная поверхность непрерывно уменьшается с ростом клетки, вследствие чего скорость роста у кокков постепенно замедляется. Удлинение клеток происходит благодаря удлинению клеточной стенки за счет включения в различные ее слои новообразованных структурных единиц. У стрептококков включение их происходит в области «экватора» клеточной стенки. У некоторых грамотрицательных бактерий этот процесс происходит без четкой локализации, т. е. в различных участках клеточной стенки. У E. coli рост наружной мембраны происходит исключительно в области ее полюсов, а рост пептидогликанового слоя – за счет включения новыхединиц в различных его участках. В условиях скоординированного роста деление клетки происходит, когда она удваивает свою биомассу, строго посередине. Процесс деления клетки сопряжен с процессом сегрегации (распределения) дочерних хромосом и дочерних плазмид в дочерние клетки. У бактерий обнаружены белкигомологи белков ParA и ParB (они осуществляют равномерное распределение плазмид между дочерними клетками) и белок MucB. Вместе с белками мембраны они образуют комплекс, растаскивающий хромосомы в дочерние клетки перед образованием межклеточной перегородки. Связь между вегетативной репликацией хромосомы и клеточным делением включает три следующих последовательных события:
1) подготовку к инициации репликации;
2) цикл репликации хромосомы (и плазмиды);
3) интервал времени между завершением репликации хромосомы и клеточным делением. Клеточный цикл бактерий можно выразить следующей формулой:
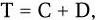
где T – время удвоения; С – время цикла репликации; D – время между завершением цикла репликации и клеточным делением.
При благоприятных условиях роста Т для E. coli и многих других бактерий составляет около 30 мин. Деление бактериальной клетки находится также под строгим генетическим контролем, нарушение которого приводит и к нарушению механизма клеточного деления.
Деление бактерий наступает в результате формирования межклеточной перегородки, которое происходит следующим образом. В том участке ЦМ, с которым связана с помощью особого рецептора молекула ДНК (хромосома, плазмида), происходят события, инициирующие процесс репликации, в результате которого вновь образующаяся дочерняя молекула ДНК прикрепляется также к рецептору на ЦМ. Область последней между двумя рецепторами, к одному из которых прикреплена родительская, а к другому – дочерняя ДНК, начинает удлиняться, в результате этого расстояние между ними все время увеличивается в течение времени С (рис. 36). По завершении процесса репликации строго по экватору между отделившимися друг от друга хромосомами начинает формироваться межклеточная перегородка путем встречной инвагинации (врастания навстречу друг к другу) ЦМ и связанной с ней области клеточной стенки (рис. 37).
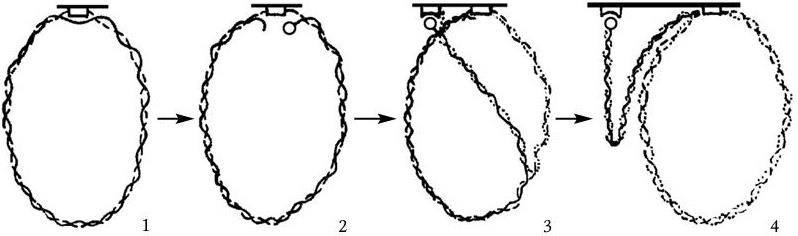
Рис. 36
Одна из нитей хромосомы прикреплена к особому рецептору мембраны (1). После инициации репликации вторая нить также разрывается (2) и прикрепляется к соседнему рецептору мембраны (3). Рост (удлинение) мембраны постепенно отделяет хромосомы друг от друга. Когда клетка удвоит свою длину, точно по ее экватору между дочерними хромосомами формируется межклеточная перегородка, и клетка делится
В результате слияния инвагинирующих участков ЦМ и КС образуется межклеточная перегородка, и родительская клетка разделяется на две дочерние клетки равной длины. Функцию аппарата митоза у бактерий выполняет ЦМ путем своего удлинения, которое раздвигает хромосомы (и плазмиды) таким образом, что они оказываются по ту и другую стороны формирующейся межклеточной перегородки в равных соотношениях.

Рис. 37
Результатов нарушения генетического контроля клеточного деления может быть по крайней мере два. Если формирования межклеточной перегородки не происходит, возникают длинные нитевидные формы. Однако при восстановлении нарушенного механизма такого контроля нити делятся на фрагменты, равные по длине нормальным клеткам. В некоторых случаях нарушение контрольных механизмов приводит к тому, что вместо одной межклеточной перегородки, формирующейся по экватору, происходит образование одной или двух перегородок, каждая из которых локализована ближе к своему полюсу. Поскольку в этом случае формирование перегородки не связано с сегрегацией хромосом, образуются так называемые мини-клетки, лишенные хромосом, которые остаются в родительской клетке. Мини-клетки могут осуществлять различные биохимические процессы, поскольку они содержат ферменты, но они не способны к размножению, так как лишены хромосом.
Помимо мини-клеток вследствие различных неблагоприятных воздействий из бактерий могут образовываться так называемые нанно-клетки, т. е. мельчайшие клетки размером 0,2 – 0,3 мкм. Их описывали под различными названиями: фильтрующиеся формы бактерий, элементарные тельца, ультрамикробактерии. Чаще всего они образуются при L-трансформации бактерий. Поскольку размеры таких клеток удобнее выражать в нанометрах, а не в долях микрометра, их стали называть нанно-клетками. Образование нанно-клеток – универсальная ответная реакция бактерий на неблагоприятные условия существования.






