Особенности роста популяции бактерий
Кинетика роста бактериальной популяции не устанавливается кинетикой роста индивидуальной клетки, хотя между ними, несомненно, существует взаимосвязь. Скорость увеличения объема индивидуальной клетки можно рассматривать как функцию времени, которое позволяет объему клетки удвоиться к концу периода между делениями. Между скоростью роста и размером клеток существуют определенные математические отношения.
Для количественной характеристики ростовых процессов в микробной популяции пользуются двумя показателями: абсолютной (валовой) скоростью и относительной (удельной) скоростью роста. Среднюю валовую скорость роста (Vср) за отрезок времени (t1 – t0) можно определить по абсолютному приросту биомассы по формуле:
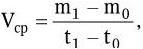
где m0 – величина биомассы в начале исследуемого отрезка времени; m1 – величина биомассы в конце исследуемого отрезка времени.
Под удельной скоростью роста (?) понимают часовой прирост, пересчитанный на единицу растущей массы:
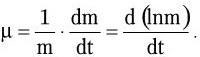
Скорость размножения бактерий ? (число удвоений за единицу времени) описывают уравнением:
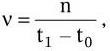
где n – число поколений.
Продолжительность жизни одного поколения (время генерации) g в среднем составляет:
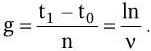
В результате логарифмирования приведенных формул и их сопоставления установлена связь удельной скорости роста с продолжительностью времени генерации и скоростью размножения клеток:
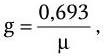
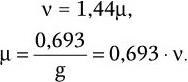
Как видно из последних формул, между временем генерации (продолжительностью жизни одного поколения) g и удельной скоростью роста ? существует обратно пропорциональная зависимость. Скорость роста микробной популяции не является величиной неизменной. В развитии микробной популяции различают следующие последовательные стадии (рис. 38): лаг-фаза; фаза положительного ускорения; фаза логарифмического роста; фаза отрицательного ускорения; стационарная фаза; фаза ускоренной гибели; фаза логарифмической гибели и фаза уменьшения скорости отмирания. Они отражают сложные процессы адаптации бактерий, привнесенных из одной среды обитания в другую, как правило, оптимальную для их размножения. Природа лаг-фазы во многом связана с тем, что в этот период происходит активный синтез всех компонентов белоксинтезирующей системы и прежде всего такого количества рибосом, которое позволило бы обеспечить максимальную активность всех биосинтетических процессов. Последующие стадии развития периодических культур отражают высокую скорость размножения бактерий. Затем, в силу постепенного истощения источника энергии и других жизненно важных метаболитов, скорость размножения бактерий уменьшается, и в стационарной фазе наступает период некоторого равновесия – количество вновь образующихся клеток становится сопоставимым с числом погибающих клеток. Вслед за этим наступает стадия, характеризующаяся постепенным уменьшением количества жизнеспособных бактерий. Это является следствием ряда причин – истощения источников энергии и других жизненно важных метаболитов, невозможности эффективно регулировать рН и rH2 среды, накопления продуктов метаболизма, тормозящих рост, и, возможно, каких-то других факторов. Очевидно, что популяция бактерий – это тоже саморегулирующаяся система, очень зависящая от среды, истощение которой оказывает на нее отрицательное действие. Жизнеспособные клетки, перенесенные из такой среды в новую питательную среду, вновь повторяют полностью весь цикл развития популяции.
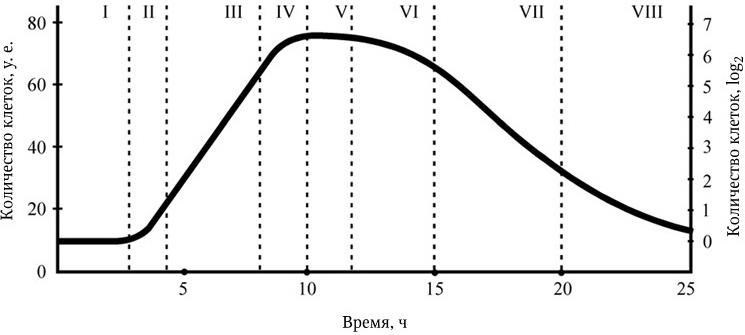
Рис. 38
I – лаг-фаза; II – фаза положительного ускорения; III – фаза логарифмического роста; IV – фаза отрицательного ускорения; V – стационарная фаза; VI – фаза ускоренной гибели; VII – фаза логарифмической гибели; VIII – фаза уменьшения скорости гибели.
На оси ординат показаны скорость размножения бактерий в условных единицах (слева) и величина популяции бактерий, выраженная логарифмом от числа живых клеток на 1 мл среды






