II.2.2 Феноптоз у растений: Снова гены смерти!
Не может быть никаких сомнений в том, что высшие организмы в том варианте их конструкции, который сегодня дошел до нас, содержат в себе семена смерти.
Быстрое старение растений, размножающихся только раз в жизни, служит, пожалуй, наиболее часто описываемым примером феноптоза, биохимический механизм которого в определенной степени уже изучен. Как пишут известный биохимик и геронтолог растений Л.Д. Нуден и его соавторы, «еще до того, как стало многое известно о биохимии старения [таких растений], оно рассматривалось как внутренне запрограммированный процесс, который специфичен и организован в отношении того, когда, где и как он происходит. Хотя это старение развивается с возрастом, оно не может быть отнесено к процессам пассивного старения, поскольку контролируется внутренними и внешними сигналами и может быть замедлено или ускорено этими сигналами. Напротив, старение как пассивное накопление повреждений, зависящее от времени, лучше всего иллюстрируется постепенным уменьшением жизнеспособности семян при их хранении» [115] (см. также [304]).
Хорошо известно, что быстрое старение и смерть сои удается предотвратить удалением стручков [184] или созревающих в них бобов [195]. Интересный опыт был поставлен Нуденом и Мюрреем [239]. На стадии созревания бобов авторы удалили у растения все стручки, кроме одного. В положенный срок (около 3 месяцев) растение не погибло, оставшись зеленым, но листья на ветке, где был оставлен единственный стручок, пожелтели и пожухли. Гибель этих листьев не удалось предотвратить, убив флоэму струей горячего пара, направленной на ветку между листьями и стручком. Пар должен был предотвратить транспорт веществ из листьев в стручок через живую ткань флоэмы, но не из стручка к листьям через ксилему, уже омертвевшую у взрослого растения. Это доказывает, что старение сои индуцируется стручками, генерирующими какой-то смертоносный сигнал или образующими ядовитое вещество, убивающее листья [238,324] (рис. II.2.2.1). Подобное наблюдение противоречит утверждению Кирквуда и Мелова, недавно опубликовавших статью о невозможности, по их мнению, существования программы старения: «Нет каких-либо указаний, — пишут авторы, — что организмы, способные к размножению только раз в жизни, активно разрушаются после завершения процесса размножения» [158].
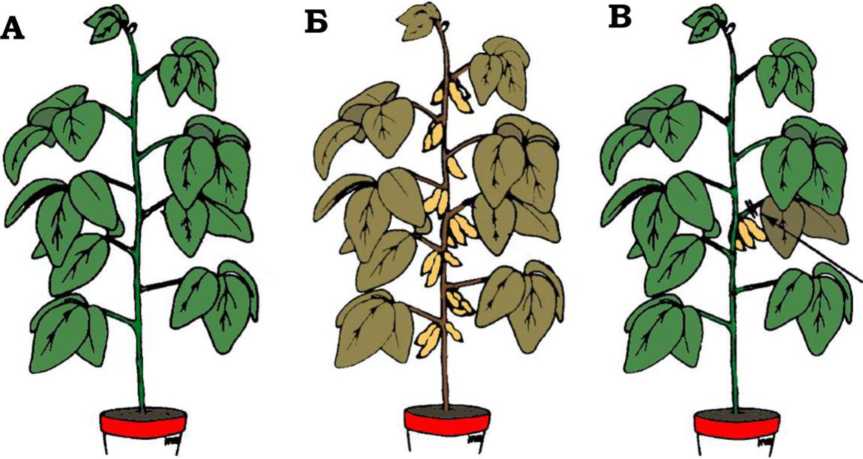
Рис. II.2.2.1
Есть основания полагать, что важную роль во включении процесса старения листьев сои играет фитогормон абсцизовая (в переводе «листопадная») кислота [66] (рис. II.2.2.2). Наномолярные концентрации этой органической кислоты регулируют экспрессию ферментов, образующих или разрушающих активные формы кислорода (АФК) в листьях растения [53]. Известно, что старение растений сопровождается резким повышением в них уровня АФК [422].
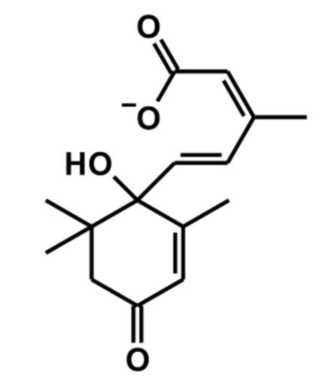
Рис. II.2.2.2
Старение сои — сравнительно быстрый процесс, занимающий примерно 10 дней при максимальном времени жизни этого растения, равном 90 дней [115]. Своевременное удаление созревающих стручков многократно увеличивает
продолжительность жизни сои. Подобный феномен описан и для других однократно размножающихся высших растений, включая резушку (Arabidopsis thaliana), классическую растительную модель биохимиков, генетиков и геронтологов (обзор см. [182]). Именно на этом объекте З. Мельцеру и сотрудникам [222,182], бельгийским биологам из Гента, удалось прямо опровергнуть еще один тезис уже упомянутой статьи Кирквуда и Мелова: «Среди многих генетических мутаций, влияющих на продолжительность жизни, иногда значительно увеличивающих эту продолжительность, еще не было найдено ни одной, которая полностью прекращала бы старение» [158]. Как сообщили Мельцер и коллеги в недавно опубликованных статьях [222,182], мутации в двух (из более чем 20 000) генов A.thaliana., а именно socl и full, имеют следствием переключение растения с полового на вегетативное размножение. Мутант зацветает с большим запозданием, с гораздо меньшим количеством цветов и семян и в то же время полностью утрачивает быстрое старение, вызываемое этими семенами. Продолжительность жизни организма (обычно менее трех месяцев) многократно увеличивается (по крайней мере до 18 месяцев). У растения появляется камбий, вторичный рост, одеревеневший ствол и множество корневищ. Из мелкой травы резушка становится кустарником с большими мясистыми листьями (рис. II.2.2.3). Цветковая меристема в основном трансформируется в вегетативную.

Рис. II.2.2.3
Авторы полагают, что современная A. thaliana исходно возникла как вегетативно размножающееся многолетнее растение, первоначально конкурировавшее с другими кустами и деревьями, и лишь затем, в процессе эволюции, стало травой (как это случилось с хвощами и папоротниками). Такой переход сопровождался появлением полового размножения, завершающегося образованием многочисленных очень мелких семян, легко разносимых ветром на большие расстояния. Семена быстро прорастают и, попав в открытую почву, дают побеги мелкой травы, которая растет, не конкурируя с другими растениями, еще не успевшими вырасти на этой почве.
Современная резушка — короткоживущий организм, убиваемый своими собственными семенами. Ранняя смерть ускоряет смену поколений, а, стало быть, и эволюцию A. thaliana. Другое обстоятельство, также ускоряющее эволюцию, состоит в том, что резушка — однократно размножающееся существо, так что новая порция семян будет заведомо образована другим индивидом, а это повысит разнообразие потомства и, стало быть, увеличит вероятность появления новых признаков. В ту же сторону (ускорения эволюции) влияет сама смена вегетативного размножения на половое, где геном потомства — всегда гибрид геномов двух родителей. Неслучайно переход к половому размножению у дрожжей есть одна из реакций на ухудшение условий среды, а значит и попытка найти новые признаки, могущие помочь выжить в изменившейся обстановке [321]. По-видимому, трансформация резушки из вегетативно размножающегося растения в цветковое произошла сравнительно недавно, так что древняя (вегетативная) программа жизни еще сохранилась в её геноме в качестве резервной[10]. A. thaliana может рассматриваться в качестве прецедента, когда инактивация нескольких генов полностью предотвращает быстрое старение, влекущее за собой смерть [324]. При этом резушка вряд ли какое-то редчайшее исключение. Мельцер и соавторы [222] пишут: «…среди покрытосеменных растений способ жизни как многолетнего дерева мог предшествовать эволюции такого растения в однолетнюю траву, причем подобные трансформации происходили неоднократно и независимо друг от друга [44]. И наоборот, однолетние травы, также многократно, эволюционировали в многолетние деревья. Например, различные виды однолетних трав, таких как Sonchus и Echium, на изолированных островах эволюционировали в древовидные многолетние растения из их континентальных однолеток [21,153,113]».
Среди многолетних растений есть примеры организмов, многие годы размножающихся вегетативно, затем переключающихся на половое размножение и гибнущих после созревания семян (рис. II.2.2.4).

Рис. II.2.2.4
Ряд видов бамбука имеет фиксированную продолжительность жизни, определяемую сроком цветения. Этот срок свой у каждого вида бамбука, варьируя от 6 лет (зацветает на шестой год жизни) до 120 лет (зацветает на сто двадцатый год). Агава (рис. II.2.2.5) и мадагаскарская пальма Ravenala madagascariensis (рис. II.2.2.6), зацветающие соответственно на десятый и сотый год, гибнут тотчас вслед за созреванием семян.

Рис. II.2.2.5

Рис. II.2.2.6
Еще дольше живет растение Puya reimodii в Андах: после 150 лет вегетативного размножения, она зацветает и вскоре гибнет. Однако даже этому долгожителю далеко до лишайников, найденных в Антарктиде и на Аляске: их возраст более 10 тыс. лет, а максимальная продолжительность вегетативной жизни просто неизвестна. На агаве был поставлен эксперимент, когда регулярное удаление цветоноса у зацветшего растения многократно продлевало его жизнь [377,421,321,228].
Во многих случаях смерть растения-родителя тотчас после созревания семян имеет очевидный биологический смысл: надо дать место потомкам. Заросли бамбука в дикой природе настолько густые, что молодые побеги не могут конкурировать с родительскими растениями, если они (родители) не освободят пространство «под солнцем» и там, где доступны влага и питательные вещества почвы [330]. Такую же роль играет смерть тропического дерева Tachigalia versicolor после созревания плодов, чтобы образовать просвет в пологе леса [259].






