Приложение 5. Предел Хейфлика, старение и теломеры
Одним из признаков старения, играющим важную роль в падении работоспособности многих органов, является снижение клеточности, т. е. уменьшение с возрастом количества функционально активных, «профильных» клеток органа. Основной гипотезой, объясняющей этот феномен, является ограниченность пролиферативного потенциала дифференцированных клеток, или «предел Хейфлика», названый так в честь Леонарда Хейфлика, элегантно и однозначно показавшего в 1961 г на человеческих фибробластах, что число делений клеток в культурах ограниченно и не превышает (для фибробластов) примерно 50 [126] (см. список литературы к приложению 5).
Природа «предела Хейфлика» оставалась загадкой до 1971 года, когда А.М. Оловников выдвинул теорию маргинотомии, согласно которой причиной остановки клеточного деления и старения в целом является следствием недорепликации последовательностей ДНК на концевых участках линейных хромосом [244]. Еще в конце 1930х годов Б. МкКлинток [220] и Г. Мёллером [[230] было отмечено, что эти концевые участки, названные теломерами (от греческого ????? — «конец» и ????? — «часть»), играют важную роль в стабилизации хромосом. Дальнейшие исследования подтвердили, что длина теломер является важнейшим параметром, влияющим на жизнь и смерть клетки.
Теломеры.
В отличие от эубактерий и архей, хранящих генетическую информацию в виде кольцевых молекул ДНК, эукариотические организмы в процессе эволюции «предпочли» линейные хромосомы. Это породило сразу две серьезные проблемы: проблему защиты концов ДНК, которые в клетке воспринимаются как двуцепочечные разрывы, подлежащие репарации, и вышеупомянутую проблему укорочения ДНК в процессе репликации, возникающей из-за неспособности ДНК-полимеразы синтезировать копию ДНК с самого начала и нуждающейся в праймере для инициации процесса синтеза. Для решения этих проблем эволюция изобрела теломеры — расположенные на концах хромосом многочисленные тандемные повторы короткой высококонсервативной последовательности нуклеотидов (TTAGGG у человека и других позвоночных), а также комплекс связанных с ними белков. При рождении длина теломерных повторов у человека составляет 10–15 тысяч пар оснований; у лабораторных мышей и крыс теломеры заметно длиннее — 20-150 тысяч пар оснований [251].
Механизм борьбы с недорепликацией был открыт в середине 1980х годов, когда обнаружилось, что в клетках одноклеточного организма тетрахимены присутствует фермент, способный наращивать теломерные участки ДНК [109]. Этот фермент, названый впоследствии теломеразой, представляет собой РНК-белковый комплекс, обладающей обратной транскриптазной активностью: для синтеза теломерной ДНК он
использует в качестве матрицы связанную с ферментом молекулу РНК, последовательность которой соответствует таковой теломерных повторов [110]. Дальнейшие исследования показали, что ген теломеразы присутствует во всех изученных эукариотических организмах.
Практически в то же время появились и первые экспериментальные подтверждений гипотезы Оловникова об укорочении теломер в процессе старения. Оказалось, что длина теломер в сперматозоидах человека заметно превышает таковую в соматических клетках взрослого организма [61]. Ряд последовавших за этим открытием экспериментов подтвердил связанное с делением клеток уменьшение длины теломер в фибробластах, клетках крови, кишечника и ряда других тканей (см. обзор [9]). Эти результаты указывали на неспособность соматических клеток человека поддерживать в процессе деления длину теломер, в отличие от клеток зародышевой линии. Также подтвердил этот вывод и тот факт, что у человека экспрессия теломеразы и теломеразная активность наблюдается только в половых клетках, клетках эмбриональной ткани и стволовых клетках [7]. (Однако у некоторых животных, в том числе у мышей, крыс [48,267,64,108], кур [369] и лягушек [24] некоторая теломеразная активность наблюдается и в соматических клетках).
Решающее значение для доказательства роли теломеразы в обеспечении неограниченного репликативного потенциала сыграли эксперименты, в которых ген этого фермента был активирован в культуре соматических клеток человека [20,368]. Оказалось, что теломеразная активность позволяла клеткам в культуре преодолеть предел Хейфлика и продолжать делиться. Скорость укорочения теломер зависит от многих факторов, и клеточное деление — лишь один из них. Здесь следует отметить, что теломеры и при делении клетки обычно укорачиваются значительно сильнее, чем можно было бы предполагать исходя только из феномена недорепликации: за одно деление соматических клеток человека теломеры уменьшаются на 20-200 нуклеотидных пар [132].
Оказалось, что важной причиной укорочения теломер является стресс, прежде всего окислительный. ДНК теломер особенно подвержена повреждениям, т. к. комплекс связанных с теломерами белков предположительно мешает доступу ферментов репарации [371,145]. Для человека показана достоверная отрицательная корреляция между уровнем стресса и длиной теломер [88]. Также было установлено, что у людей регулярная физическая нагрузка коррелирует с более медленным укорочением теломер [174,247].
Сокращение длины теломер ниже определенного предела приводит к остановке клеточного деления. Происходит это, однако, не из-за постепенного «съедания» кодирующей области ДНК вследствие недорепликации. Минимальная длина теломер в клетках человеческого организма составляет около 500 пар оснований [7]. Показано также, что длина теломер, начиная с которой дальнейшее укорочение приводит к нестабильности генома, слиянию хромосом и иным аномалиям, составляет примерно 13 теломерных повторов (около 100 пар оснований) [42].
Механизм остановки клеточного деления при достижении определенной, минимальной длины теломер до конца не ясен, и включает много факторов, среди которых важнейшую роль играет «страж генома» — белок p53, а также белки p21, p16INK4A и pRb (рис. П-5.1) [306]. Все они являются ключевыми регуляторами клеточного ответа на разные виды стресса (в том числе на повреждение ДНК) и в защите от трансформации клетки в раковую.
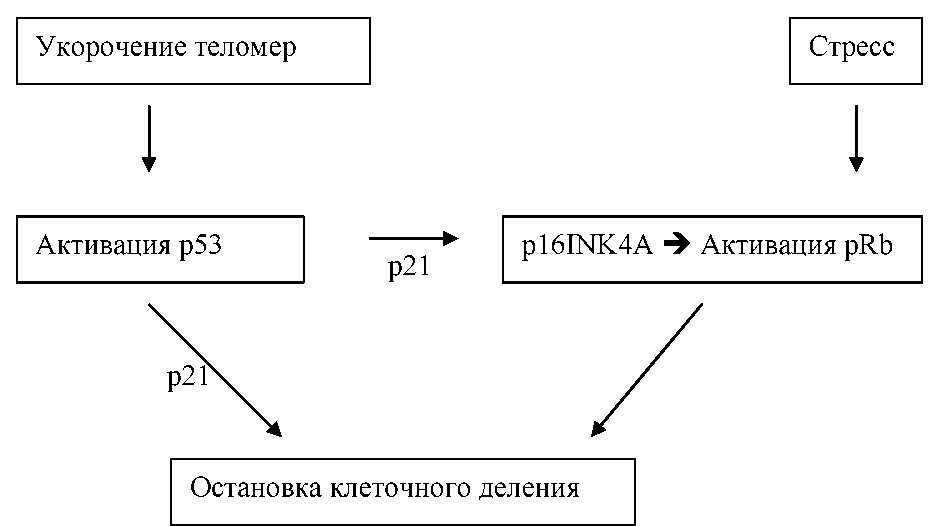
Рис. II-5.1.
Ограничение клеточного деления, не допускающее бесконтрольную пролиферацию и рак, является, по-видимому, одной из основных функций сокращения теломер в соматических клетках in vivo [7]. На это указывает экспрессия теломеразы и присутствие теломеразной активности в клетках более чем 90 % раковых опухолей (см. также обзор [307]). Эксперименты на мышах выявили, что потеря теломеразной активности (при условии нормального уровня активности белка p53) повышала устойчивость животных к раковым заболеваниям [216]. Существенно, что вероятность возникновения рака в пересчете на одну
клетку у мышей и крыс (экспрессирующих теломеразу в соматических клетках) на порядки выше, чем у человека.
Старение.
Вскоре после открытия предела Хейфлика появились теории «клеточного старения», объяснявшие феномен падения жизненных функций организма с возрастом снижением репликативного потенциала его клеток. Исследования теломер и теломеразы дали объяснение феномену предела Хейфлика, что вызвало рост интереса к этому ферменту со стороны медиков и геронтологов. Было выяснено, что у человека дефект даже в одном из аллелей генов теломеразы приводит к патологиям, из которых наиболее распространен врожденный дискератоз. Причиной этого вероятно является жесткая регуляция экспрессии теломеразы (как ее белкового, так и РНК-компонента) [7].
Кроме того, была выявлена корреляция между чрезмерно укороченными теломерами и частотой ряда возраст-зависимых заболеваний [7]. Одной из наиболее ярких работ по роли теломеразы в процессе старения стали эксперименты, в которых с помощью суперэкспрессии теломеразы была увеличена продолжительность жизни мышей, при условии одновременного усиления антираковой защиты [357]. Следует отметить, что наличие компромисса между защитой от рака и замедлением старения есть не только в случае теломеразы. Другим известным примером является белок p53, который с одной стороны является ключевым звеном антираковой защиты, а с другой — ускоряет старение [365].
Не менее удивительные результаты были получены в группе Марии Бласко: исследователям удалось замедлить старение и продлить жизнь у взрослых мышей с помощью повышения уровня экспрессии теломеразы [15]. Немаловажно, что в этой работе «теломеразная генная терапия», осуществлявшаяся с помощью обработки животных аденоассоциированным вирусом, оказалась эффективной не только для взрослых (1 год), но и для старых (2 года) мышей: медианная продолжительность жизни после обработки вирусов возросла на 24 % и на 13 % соответственно. Примечательно, что при этом терапия не привела к увеличению частоты возникновения злокачественных образований [15].
Но, несмотря на всю привлекательность гипотезы о неуклонно сокращающихся теломерах как молекулярных часах клетки, многие ее аспекты остаются неясными, а ряд фактов вступает с ней в противоречие. Как уже было отмечено выше, у мышей и крыс теломеры почти на порядок длиннее, чем у человека, а теломераза активна и в соматических клетках. Однако стареют эти грызуны значительно быстрее людей. Эксперименты на мышах, у которых был удален ген теломеразы, показали, что животные жизнеспособны и плодовиты в течение 6 поколений [18]. Скорость укорочения теломер у мутантных животных составила 4.8±2.4 тысячи пар оснований на поколение, что сравнимо со скоростью укорочения теломер у человека. В культуре клеток из мутантных животных в более поздних поколениях наблюдался рост частоты хромосомных аберраций, однако влияния укорочения теломер на значение предела Хейфлика не было выявлено. Этот факт, а также ряд других наблюдений [7] указывают, что укорочение теломер является далеко не единственным механизмом ограничения пролиферации соматических клеток.
Следует также отметить, что экспериментальные доказательства роли феномена предела Хейфлика в старении многоклеточных организмов достаточно немногочисленны, и интерпретация данных, полученных на культурах, где клетки подвергаются ряду нефизиологических стрессовых факторов, требует осторожности [72]. В частности, показано, что снижение концентрации кислорода от обычного значения 20 % до 1 % приводило к замедлению репликативного старения фибробластов человека [293], а также что концентрация кислорода ограничивает пролиферацию в культуре мышиных фибробластов [256].
Однако теломеры могут играть важную роль в процессе старения и вне контекста ограничения пролиферативной активности. Как было отмечено выше, теломерные участки ДНК хромосом более уязвимы для различных вредоносных стрессовых факторов, в том числе АФК. Таким образом, теломеры могут служить индикатором стресса в клетке, генерируя сигнал «повреждение ДНК!» при относительно низком уровне стресса, еще не слишком опасном для остальной хромосомной ДНК.
Заключение
Укорочение теломер является важным (но не единственным!) физиологическим механизмом, задействованным в ограничении пролиферации соматических клеток и предотвращении возникновения рака. Весьма вероятно, что этот феномен приводит также к сокращению с возрастом числа функциональных клеток в ряде органов, и — как следствие — к снижению жизнеспособности организма, т. е. к старению.
Интересным вопросом, связанным с эволюционным значением теломер, является причина перехода от кольцевой ДНК к линейной. Согласно гипотезе Боулза [25], этот переход — ни что иное, как возникновение первой «программы старения» у ранних, одноклеточных форм жизни: неуклонное укорачивание ДНК при каждом следующем делении приводит в итоге к падению жизнеспособности, и тем самым ускоряет смену поколений.
Однако универсальность данного механизма старения представляется маловероятной. Об этом свидетельствует отсутствие ускоренного старения у мышей, лишенных теломеразы. Еще один пример: у нематоды Caenorhabditis elegans вообще отсутствует деление соматических клеток во взрослом состоянии, что никак не мешает прогрессирующему старению этого хорошо изученного модельного организма.
К сожалению, наши знания о роли теломер в жизни и смерти клеток пока недостаточны для построения полной непротиворечивой картины. Поэтому исследование теломер остается одной из интереснейших и многообещающих областей клеточной и молекулярной биологии.






