II.6.6.2 Запрограммированное старение организма — многостадийный процесс
Ввиду неясности природы ювенильного гормона на сегодня остается открытым вопрос, каким именно образом этот гормон сдерживает нарастание уровня АФК в митохондриях. Вряд ли мы имеем здесь дело с прямым антиоксидантным действием гормона. Любой гормон занимает слишком высокое положение в иерархии управленческих устройств организма, чтобы “лично” участвовать в регуляции того или иного процесса. Скорее речь должна идти об еще одном белке — рецепторе, который связывает ювенильный гормон и множит передаваемый им сигнал.
Еще более неопределенной остается проблема передачи сигнала старения внутри клетки периферической ткани — от клеточной поверхности, непосредственно взаимодействующей с ювенильным гормоном, к митохондриям. Это может быть, например, сам ювенильный гормон, если он проникает в клетку и достигает митохондрий. Есть указания, что, например, мелатонин концентрируется в митохондриях [102,201]. Однако неясно, как влияет мелатонин на уровень митохондриальных АФК. Это влияние, если и есть, вряд ли сводится к его способности действовать в качестве антиоксиданта: мелатонина просто не хватит, чтобы вызвать измеримое снижение АФК. Значит, надо думать о каком-то рецепторе мелатонина в митохондриях, передающем сигнал к снижению уровня АФК[23]. Такое снижение может быть результатом замедления образования митохондриальных АФК либо ускорения их обезвреживания. В пользу первого варианта говорит тот факт, что обнаружена отрицательная корреляция продолжительности жизни и скорости генерации АФК при обратном переносе электронов через комплекс I дыхательной цепи (см. выше рис. II.6.5.2). По нашим данным [318,164,320] и результатам Л. Шу-сена [198,197], генерация АФК при обратном переносе электронов пороговым образом зависит от величины мембранного потенциала (??) на митохондриях. Так, снижение ?? на 15 % ведет к десятикратному уменьшению генерации АФК, причем эффект не зависит от того, каким именно образом достигнуто снижение ??: добавлением АДФ (то есть включением окислительного фосфорилирования), малой дозой разобщителя-протонофора или ингибитором дыхания. Пороговая зависимость генерации АФК от ?? была впоследствии подтверждена в других лабораториях [372,342]. Основываясь на этих наблюдениях, мы можем предположить, что при старении происходит повышение ?? в митохондриях, и именно это обстоятельство увеличивает генерацию АФК при обратном переносе электронов[24]. В свою очередь, повышение ?? может быть результатом уменьшения утечки Н+ через мембрану митохондрий. Этот процесс катализируется разобщающими белками и митохондриальными переносчиками анионов, облегчающими электрофорез анионов жирных кислот, то есть их движение изнутри митохондрий наружу (см. обзор [320]). Снаружи анионы жирных кислот протонируются и возвращаются назад в митохондриальный матрикс, неся протоны (разобщающий «жирнокислотный цикл» [317]). Активность жирнокислотного цикла может регулироваться либо белками-переносчиками анионов жирных кислот, либо концентрацией этих кислот в цитозоле. Соответственно, рост ?? при старении может быть обусловлен либо снижением внутриклеточного уровня свободных жирных кислот, либо уменьшением активности белков-переносчиков (например, если предположить, что мелатонин как-то активирует перенос анионов жирных кислот этими белками).
В нашей лаборатории было продемонстрировано, как повышение ?? митохондрий участвует в быстром феноптозе дрожжей в ответ на амиодарон или избыток феромона [266]. Была выявлена следующая цепь событий:
индуктор феноптоза ? ??? ? АФК? ? ??? ? гибель клетки.
Интересно, что в норме дрожжи, выращенные на углеводной среде, имели низкий ?? и слабое дыхание, которое не ингибировалось олигомицином и не стимулировалось небольшими дозами разобщителя. Добавление амиодарона или феромона повышало ??, стимулировало дыхание и вызывало появление чувствительности дыхания к малым дозам разобщителя. Феноптоз мог быть блокирован низкими концентрациями разобщителя, резко повышавшего выживаемость дрожжей в присутствии амиодарона или феромона. Существенно, что первоначальный резкий подъем генерации АФК при дыхании под действием повышенного ?? может быть временным. Он играет роль запала в лавинообразном процессе (цепной реакции) перекисного окисления липидов митохондрий. Как только количество цитохрома с, вышедшего в межмембранное пространство, и окисленного кардиолипина, активирующего перекисное окисление этого фосфолипида цитохромом с, окажутся достаточными, наступит новый этап образования АФК митохондриями, потерявшими свои главные механизмы антиоксидантной защиты из-за резкого повышения проводимости внутренней митохондриальной мембраны вследствие окисления кардиолипина. Повышение проводимости ведет к падению ??, выключению энергозависимой трансгидрогеназы, окислению NADPH и глутатиона и выходу последнего из матрикса митохондрий в цитозоль.
Дальнейшие этапы старческого феноптоза, указанные на нашей схеме (рис. II.6.6.1), не требуют подробных комментариев. Общеизвестно, что возрастание уровня АФК в митохондриях приводит к открытию неспецифической поры во внутренней мембране этих органелл, что, в свою очередь, ведет к набуханию митохондриального матрикса, разрыву внешней мембраны митохондрий и выходу в цитозоль проапоптических белков (типа цитохрома с), в норме спрятанных в межмембраннике митохондрий. Так активируется апоптоз [319,245,248].
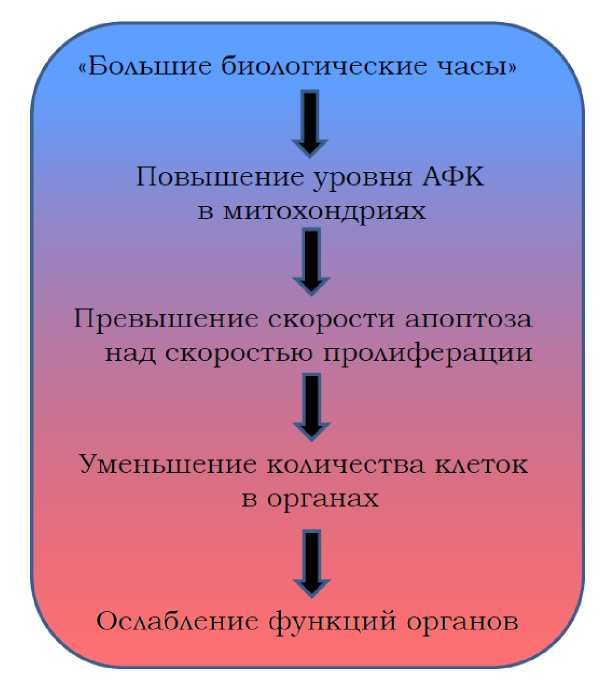
Рис. II.6.6.1
С возрастом количество клеток, уходящих в апоптоз, становится столь велико, что не восполняется появлением новых клеток. Общее число клеток в органах уменьшается, а вместе с ним падает способность этих органов совершать полезную работу, то есть происходит старение организма [435]. (О возможной роли теломер в старении см. Приложение 5).