Центральная нервная система
СПИННОЙ МОЗГ
Развитие спинного мозга. Как уже отмечалось, филогенетически спинной мозг (туловищный мозг ланцетника) появляется на III этапе развития нервной системы (трубчатая нервная система). В это время головного мозга еще нет, поэтому туловищный мозг имеет центры для управления всеми процессами организма, как вегетативными, так и анимальными (висцеральные и соматические центры). Соответственно сегментарному строению тела туловищный мозг имеет сегментарное строение, он состоит из связанных между собой невромеров, в пределах которых замыкается простейшая рефлекторная дуга. Метамерное строение спинного мозга сохраняется и у человека, чем и обусловливается наличие у него коротких рефлекторных дуг.
С появлением головного мозга (этап кефализации) в нем возникают высшие центры управления всем организмом, а спинной мозг попадает в подчиненное положение. Спинной мозг не остается только сегментарным аппаратом, а становится и проводником импульсов от периферии к головному мозгу и обратно, в нем развиваются двусторонние связи с головным мозгом. Таким образом, в процессе эволюции спинного мозга образуется два аппарата: более старый сегментарный аппарат собственных связей спинного мозга и более новый надсегменгарный аппарат двусторонних проводящих путей к головному мозгу. Такой принцип строения наблюдается и у человека.
Решающим фактором образования туловищного мозга является приспособление к окружающей среде при помощи движения. Поэтому строение спинного мозга отражает способ передвижения животного. Так, например, у пресмыкающихся, не имеющих конечностей и передвигающихся с помощью туловища (например, у змеи), спинной мозг развит равномерно на всем протяжении и не имеет утолщений. У животных, пользующихся конечностями, возникает два утолщения, при этом, если более развиты передние конечности (например, крылья птиц), то преобладает переднее (шейное) утолщение спинного мозга; если более развиты задние конечности (например, ноги страуса), то увеличено заднее (поясничное) утолщение; если в ходьбе участвуют и передние, и задние конечности (четвероногие млекопитающие), то одинаково развиты оба утолщения. У человека в связи с более сложной деятельностью руки как органа труда шейное утолщение спинного мозга дифференцировалось сильнее, чем поясничное.
Отмеченные факторы филогенеза играют роль в развитии спинного мозга и в онтогенезе. Спинной мозг развивается из нервной трубки, из ее заднего отрезка (из переднего возникает головной мозг). Из вентрального отдела трубки образуются передние столбы серого вещества спинного мозга (клеточные тела двигательных нейронов), прилегающие к ним пучки нервных волокон и отростки названных нейронов (двигательные корешки). Из дорсального отдела возникают задние столбы серого вещества (клеточные тела вставочных нейронов), задние канатики (отростки чувствительных нейронов).
Таким образом, вентральная часть мозговой трубки является первично двигательной, а дорсальная — первично чувствительной. Деление на моторную (двигательную) и сенсорную (чувствительную) области простирается на всю нервную трубку и сохраняется в стволе головного мозга.
Из-за редукции каудальной части спинного мозга получается тонкий тяж из нервной ткани, будущая f?lum termin?le. Первоначально, на 3-м месяце утробной жизни, спинной мозг занимает весь позвоночный канал, затем позвоночник начинает расти скорее, чем мозг, вследствие чего конец последнего постепенно перемещается кверху (краниально). При рождении конец спинного мозга уже находится на уровне III поясничного позвонка, а у взрослого достигает высоты I–II поясничного позвонка. Благодаря такому «восхождению» спинного мозга отходящие от него нервные корешки принимают косое направление (рис. 267).
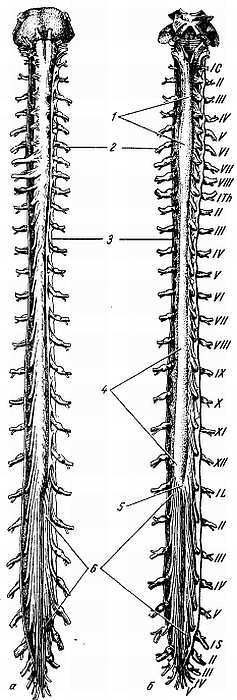
Рис. 267.
а — вид спереди; б — вид сзади. Твердая и паутинная оболочки разрезаны. Сосудистая оболочка снята. Римскими цифрами обозначен порядок расположения шейных (С), грудных (Th), поясничных (L) и крестцовых (S) спинномозговых нервов; 1 — intumescentia cervicalis; 2 — gangl. spinale; 3 — dura mater medullae spinalis; 4 — intumescentia lumbosacralis; 5 — conus medullaris; 6 — cauda equina.
Строение спинного мозга
Спинной мозг, med?lla spin?lis (греч. myel?s), лежит в позвоночном канале и у взрослых представляет собой длинный (45 см у мужчин и 41–42 см у женщин), несколько сплюснутый спереди назад цилиндрический тяж, который вверху (краниально) непосредственно переходит в продолговатый мозг, а внизу (каудально) оканчивается коническим заострением, c?nus medull?ris, на уровне II поясничного позвонка (см. рис. 267). Знание этого факта имеет практическое значение (чтобы не повредить спинной мозг при поясничном проколе с целью взятия спинномозговой жидкости или с целью спинномозговой анестезии, надо вводить иглу шприца между остистыми отростками III и IV поясничных позвонков).
От c?nus medull?ris отходит книзу так называемая концевая нить, f?lum termin?le, представляющая атрофированную нижнюю часть спинного мозга, которая внизу состоит из продолжения оболочек спинного мозга и прикрепляется ко II копчиковому позвонку.
Спинной мозг на своем протяжении имеет два утолщения, соответствующих корешкам нервов верхней и нижней конечностей: верхнее из них называется шейным утолщением, intumesc?ntia cervic?lis, а нижнее — пояснично-крестцовым, intumesc?ntia lumbosacr?lis. Из этих утолщений более обширно пояснично-крестцовое, но более дифференцировано шейное, что связано с более сложной иннервацией руки как органа труда. Образовавшимися вследствие утолщения боковых стенок спинномозговой трубки и проходящими по средней линии передней и задней продольными бороздами: глубокой f?ssura medi?na ant?rior, и поверхностной, s?lcus medi?nus post?rior, спинной мозг делится на две симметричные половины — правую и левую; каждая из них в свою очередь имеет слабо выраженную продольную борозду, идущую по линии входа задних корешков (s?lcus posterolater?lis) и по линии выхода передних корешков (s?lcus anterolater?lis).
Эти борозды делят каждую половину белого вещества спинного мозга на три продольных канатика: передний — fun?culus ant?rior, боковой — fun?culus later?lis и задний — fun?culus post?rior. Задний канатик в шейном и верхнегрудном отделах делится еще промежуточной бороздкой, s?lcus interm?dius post?rior, на два пучка: fasc?culus gr?cilis и fasc?culus cune?tus. Оба эти пучка под теми же названиями переходят вверху на заднюю сторону продолговатого мозга.
На той и другой стороне из спинного мозга выходят двумя продольными рядами корешки спинномозговых нервов. Передний корешок, r?dix ventr?lis s. ant?rior, выходящий через s?lcus anterolater?lis, состоит из нейритов двигательных (центробежных, или эфферентных) нейронов, клеточные тела которых лежат в спинном мозге, тогда как задний корешок, r?dix dors?lis s. post?rior, входящий в s?lcus posterolateralis, содержит отростки чувствительных (центростремительных, или афферентных) нейронов, тела которых лежат в спинномозговых узлах.
На некотором расстоянии от спинного мозга двигательный корешок прилегает к чувствительному (рис. 268) и они вместе образуют ствол спинно-мозгового нерва, tr?ncus n. spin?lis, который невропатологи выделяют под именем канатика, fun?culus.
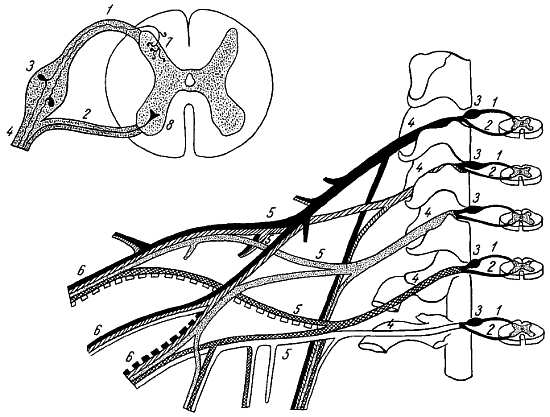
Рис. 268.
1 — radix posterior; 2 — radix anterior; 3 — gangl. spinale; 4 — truncus n. spinalis; 5 — plexus; 6 — ветви сплетения; 7 — задний рог; 8 — передний рог.
При воспалении канатика (фуникулит) возникают сегментарные расстройства одновременно двигательной и чувствительной сфер; при заболевании корешка (радикулит) наблюдаются сегментарные нарушения одной сферы — или чувствительной, или двигательной, а при воспалении ветвей нерва (неврит) расстройства соответствуют зоне распространения данного нерва. Ствол нерва обычно очень короткий, так как по выходе из межпозвоночного отверстия нерв распадается на свои основные ветви.
В межпозвоночных отверстиях вблизи места соединения обоих корешков задний корешок имеет утолщение — спинномозговой узел, g?nglion spin?le, содержащий ложноуниполярные нервные клетки (афферентные нейроны) с одним отростком, который делится затем на две ветви: одна из них, центральная, идет в составе заднего корешка в спинной мозг, другая, периферическая, продолжается в спинномозговой нерв. Таким образом, в спинномозговых узлах отсутствуют синапсы, так как здесь лежат клеточные тела только афферентных нейронов. Этим названные узлы отличаются от вегетативных узлов периферической нервной системы, так как в последних вступают в контакты вставочные и эфферентные нейроны. Спинномозговые узлы крестцовых корешков лежат внутри крестцового канала, а узел копчикового корешка — внутри мешка твердой оболочки спинного мозга.
Вследствие того что спинной мозг короче позвоночного канала, место выхода нервных корешков не соответствует уровню межпозвоночных отверстий. Чтобы попасть в последние, корешки направляются не только в стороны от мозга, но еще и вниз, при этом тем отвеснее, чем ниже они отходят от спинного мозга. В поясничной части последнего нервные корешки спускаются к соответствующим межпозвоночным отверстиям параллельно f?lum termin?le, облекая ее и c?nus medull?ris густым пучком, который носит название конского хвоста, cauda equina (см. рис. 267).
Внутреннее строение спинного мозга. Спинной мозг состоит из серого вещества, содержащего нервные клетки, и белого вещества, слагающегося из миелиновых нервных волокон.
А. Серое вещество, subst?ntia gr?sea, заложено внутри спинного мозга и окружено со всех сторон белым веществом. Серое вещество образует две вертикальные колонны, помещенные в правой и левой половинах спинного мозга. В середине его заложен узкий центральный канал, can?lis centr?lis, спинного мозга, проходящий во всю длину последнего и содержащий спинномозговую жидкость. Центральный канал является остатком полости первичной нервной трубки. Поэтому вверху он сообщается с IV желудочком головного мозга, а в области c?nus medull?ris заканчивается расширением — концевым желудочком, ventr?culus tcrmin?lis.
Серое вещество, окружающее центральный канал, носит название промежуточного, subst?ntia interm?dia centr?lis. В каждой колонне серого вещества два столба: передний, col?mna ant?rior, и задний, col?mna post?rior.
На поперечных разрезах спинного мозга эти столбы имеют вид рогов: переднего, расширенного, c?rnu ant?rius, и заднего, заостренного, c?rnu post?rius. Поэтому общий вид серого вещества на фоне белого напоминает букву «Н».
Серое вещество состоит из нервных клеток, группирующихся в ядра, расположение которых в основном соответствует сегментарному строению спинного мозга и его первичной трехчленной рефлекторной дуге. Первый, чувствительный, нейрон этой дуги лежит в спинномозговых узлах, периферический отросток которого начинается рецепторами в органах и тканях, а центральный в составе задних чувствительных корешков проникает через s?lcus posterolater?lis в спинной мозг. Вокруг верхушки заднего рога образуется пограничная зона белого вещества, представляющая собой совокупность центральных отростков клеток спинномозговых узлов, заканчивающихся в спинном мозге. Клетки задних рогов образуют отдельные группы или ядра, воспринимающие из сомы различные виды чувствительности, — соматически-чувствительные ядра. Среди них выделяются: грудное ядро, n?cleus thor?cicus (col?mna thor?cica), наиболее выраженное в грудных сегментах мозга; находящееся на верхушке рога студенистое вещество, subst?ntia gelatin?sa, а также так называемые собственные ядра, n?clei pr?prii.
Заложенные в заднем роге клетки образуют вторые, вставочные, нейроны.
В сером веществе задних рогов разбросаны также рассеянные клетки, так называемые пучковые клетки, аксоны которых проходят в белом веществе обособленными пучками волокон. Эти волокна несут нервные импульсы от определенных ядер спинного мозга в его другие сегменты или служат для связи с третьими нейронами рефлекторной дуги, заложенными в передних рогах того же сегмента. Отростки этих клеток, идущие от задних рогов к передним, располагаются вблизи серого вещества, по его периферии, образуя узкую кайму белого вещества, окружающего серое со всех сторон. Это собственные пучки спинного мозга, fasciculi pr?prii. Вследствие этого раздражение, идущее из определенной области тела, может передаваться не только на соответствующий ей сегмент спинного мозга, но захватывать и другие. В результате простой рефлекс может вовлекать в ответную реакцию целую группу мышц, обеспечивая сложное координированное движение, остающееся, однако, безусловнорефлекторным.
Передние рога содержат третьи, двигательные, нейроны, аксоны которых, выходя из спинного мозга, составляют передние, двигательные, корешки. Эти клетки образуют ядра эфферентных соматических нервов, иннервирующих скелетную мускулатуру, — сомагически-двигательные ядра. Последние имеют вид коротких колонок и лежат в виде двух групп — медиальной и латеральной. Нейроны медиальной группы иннервируют мышцы, развившиеся из дорсальной части миотомов (аутохтонная мускулатура спины), а латеральной — мышцы, происходящие из вентральной части миотомов (вентролатеральные мышцы туловища и мышцы конечностей); чем дистальнее иннервируемые мышцы, тем латеральнее лежат иннервирующие их клетки.
Наибольшее число ядер содержится в передних рогах шейного утолщения спинного мозга, откуда иннервируются верхние конечности, что определяется участием последних в трудовой деятельности человека. У последнего в связи с усложнением движений руки как органа труда этих ядер значительно больше, чем у животных, включая антропоидов. Таким образом, задние и передние рога серого вещества имеют отношение к иннервации органов животной жизни, особенно аппарата движения, в связи с усовершенствованием которого в процессе эволюции и развивался спинной мозг.
Передний и задний рога в каждой половине спинного мозга связаны между собой промежуточной зоной серого вещества, которая в грудном и поясничном отделах спинного мозга, на протяжении от I грудного до II–III поясничных сегментов особенно выражена и выступает в виде бокового рога, c?rnu later?le. Вследствие этого в названных отделах серое вещество на поперечном разрезе приобретает вид бабочки. В боковых рогах заложены клетки, иннервирующие вегетативные органы и группирующиеся в ядро, которое носит название col?mna intermediolater?lis. Нейриты клеток этого ядра выходят из спинного мозга в составе передних корешков.
Б. Белое вещество, subst?ntia ?lba, спинного мозга состоит из нервных отростков, которые составляют три системы нервных волокон:
1. Короткие пучки ассоциативных волокон, соединяющих участки спинного мозга на различных уровнях (афферентные и вставочные нейроны). 2. Длинные центростремительные (чувствительные, афферентные). 3. Длинные центробежные (двигательные, эфферентные). Первая система (коротких волокон) относится к собственному аппарату спинного мозга, а остальные две (длинных волокон) составляют проводниковый аппарат двусторонних связей с головным мозгом.
Собственный аппарат включает серое вещество спинного мозга с задними и передними корешками и собственными пучками белого вещества (fasc?culi pr?prii), окаймляющими серое в виде узкой полосы. По развитию собственный аппарат является образованием филогенетически более старым и потому сохраняет примитивные черты строения — сегментарносгь, отчего его называют также сегментарным аппаратом спинного мозга в отличие от остального несегментированного аппарата двусторонних связей с головным мозгом.
Таким образом, нервный сегмент — это поперечный отрезок спинного мозга и связанных с ним правого и левого спинномозговых нервов, развившихся из одного невротома (невромера). Он состоит из горизонтального слоя белого и серого вещества (задние, передние и боковые рога), содержащего нейроны, отростки которых проходят в одном парном (правом и левом) спинномозговом нерве и его корешках (см. рис. 268). В спинном мозге различают 31 сегмент, которые топографически делятся на 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 копчиковый. В пределах нервного сегмента замыкается короткая рефлекторная дуга (см. рис. 264).
Так как собственный сегментарный аппарат спинного мозга возник тогда, когда еще не было головного, то функция его — это осуществление тех реакций в ответ на внешнее и внутреннее раздражения, которые в процессе эволюции возникли раньше, т. е. врожденных реакций.
Аппарат двусторонних связей с головным мозгом филогенетически более молодой, так как возник лишь тогда, когда появился головной мозг.
По мере развития последнего разрастались кнаружи и проводящие пути, связывающие спинной мозг с головным (рис. 269).
Рис. 269.
Этим объясняется тот факт, что белое вещество спинного мозга как бы окружило со всех сторон серое вещество. Благодаря проводниковому аппарату собственный аппарат спинного мозга связан с аппаратом головного мозга, который объединяет работу всей нервной системы. Нервные волокна группируются в пучки, а из пучков составляются видимые невооруженным глазом канатики: задний, боковой и передний. В заднем канатике (рис. 270), прилежащем к заднему (чувствительному) рогу, лежат пучки восходящих нервных волокон; в переднем канатике, прилежащем к переднему (двигательному) рогу, лежат пучки нисходящих нервных волокон; наконец, в боковом канатике находятся и те и другие. Кроме канатиков, белое вещество находится в белой спайке, comiss?ra ?lba, образующейся вследствие перекреста волокон спереди от subst?ntia interm?dia centr?lis; сзади белая спайка отсутствует.
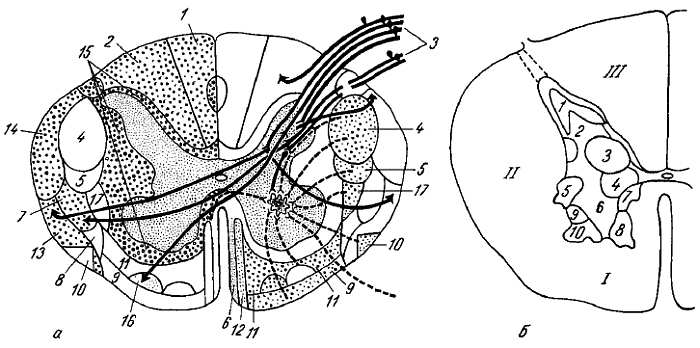
Рис. 270.
а — схема проводящих путей спинного мозга: слева изображено местоположение восходящих, справа — нисходящих систем волокон: 1 — fasc. gracilis; 2 — fasc. cuneatus; 3 — radix posterior; 4 — tr. corticospinalts lateralis; 5 — tr. rubrospinalis; 6 — tr. tectospinalis; 7 — tr. spinothalamicus lateralis; 8 — tr. spinotectalis; 9 — tr. vestibulospinalis; 10 — tr. olivospinalis; 11 — tr. reticulospinalis; 12 — tr. corticospinalis anterior; 13 — tr. spinocerebellaris anterior; 14 — tr. spinocerebellaris posterior; 15 — fascc. proprii; 16 — tr. spinothalamicus anterior; 17 — tr. thalamospinalis;
б — ядра серого вещества (в грудном отделе): 1 — substantia gelatinosa; 2 — nucl. proprius cornu posterioris; 3 — nuc{. thoracicus; 4 — nucl intermediomedialis; 5 — columna intermediolateralis; 6, 7, 8, 9, 10 — пять двигательных ядер переднего рога;
I, II, III — соответственно передний, боковой и задний канатики белого вещества.
Задние канатики содержат волокна задних корешков спинномозговых нервов, слагающиеся в две системы:
1. Медиально расположенный тонкий пучок, fasc?culus gr?cilis.
2. Латерально расположенный клиновидный пучок, fasc?culus cune?tus.
Пучки тонкий и клиновидный проводят от соответствующих частей тела к коре головного мозга сознательную проприоцептивную (мышечно-суставное чувство) и кожную (чувство стереогноза — узнавание предметов на ощупь) чувствительность, имеющую отношение к определению положения тела в пространстве, а также тактильную чувствительность.
Боковые канатики содержат следующие пучки:
А. Восходящие.
К заднему мозгу: 1) tr?ctus spinocerebell?ris post?rior, задний спинно-мозжечковый путь, располагается в задней часта бокового канатика по его периферии; 2) tr?ctus spinocerebell?ris ant?rior, передний спинно-мозжечковый путь, лежит вентральнее предыдущего.
Оба спинно-мозжечковых тракта проводят бессознательные проприоцеитивные импульсы (бессознательная координация движений).
К среднему мозгу: 3) tr?ctus spinotect?lis, спинно-покрышечный путь, прилегает к медиальной стороне и передней части tr?ctus spinocerebell?ris ant?rior.
К промежуточному мозгу: 4) tr?ctus spinothal?micus later?lis прилегает с медиальной стороны к tr?ctus spinocerebell?ris ant?rior, тотчас позади tr?ctus spinotect?lis. Он проводит в дорсальной части тракта температурные раздражения, а в вентральной — болевые; 5) tr?ctus spinothal?micus ant?riror s. ventr?lis аналогичен предыдущему, но располагается кпереди от соименного латерального и является путем проведения импульсов осязания, прикосновения (тактильная чувствительность). По последним данным, этот тракт располагается в переднем канатике.
Б. Нисходящие.
От коры большого мозга: 1) латеральный корково-спинномозговой (пирамидный) путь, tr?ctus corticospin?lis (pyramid?lis) later?lis. Этот тракт является сознательным эфферентным двигательным путем.
От среднего мозга: 2) tr?ctus rubrospin?lis. Он является бессознательным эфферентным двигательным путем.
От заднего мозга: 3) tr?ctus olivospin?lis, лежит вентральнее tr?ctus spinocerebell?ris anterior, вблизи переднего канатика.
Передние канатики содержат нисходящие пути.
От коры головного мозга: 1) передний корково-спинномозговой (пирамидный) путь, tr?ctus corticospin?lis (pyramid?lis) ant?rior, составляет с латеральным пирамидным пучком общую пирамидную систему.
От среднего мозга: 2) tr?ctus tectospin?lis, лежит медиальнее пирамидного пучка, ограничивая fiss?ra medi?na ant?rior. Благодаря ему осуществляются рефлекторные защитные движения при зрительных и слуховых раздражениях — зрительно-слуховой рефлекторный тракт.
Ряд пучков идет к передним рогам спинного мозга от различных ядер продолговатого мозга, имеющих отношение к равновесию и координации движений, а именно:
3) от ядер вестибулярного нерва — tr?ctus vestibulospin?lis — лежит на границе переднего и бокового канатиков;
4) от form?tio reticul?ris — tr?ctus reticulospin?lis ant?rior, лежит в средней части переднего канатика;
5) собственно пучки, fasc?culi pr?prii, непосредственно прилегают к серому веществу и относятся к собственному аппарату спинного мозга.
Оболочки спинного мозга
Спинной мозг одет тремя соединительноткаными оболочками, men?nges, происходящими из мезодермы. Оболочки эти следующие, если идти с поверхности вглубь: твердая оболочка, d?ra m?ter; паутинная оболочка, arachno?dea, и мягкая оболочка, pia m?ter. Краниально все три оболочки продолжаются в такие же оболочки головного мозга.
1. Твердая оболочка спинного мозга, d?ra m?ter spin?lis, облекает в форме мешка снаружи спинной мозг. Она не прилегает вплотную к стенкам позвоночного канала, которые покрыты надкостницей. Последнюю называют также наружным листком твердой оболочки. Между надкостницей и твердой оболочкой находится эпидуральное пространство, с?vitas epidur?lis. В нем залегают жировая клетчатка и венозные сплетения — pl?xus ven?si vertebr?les int?rni, в которые вливается венозная кровь от спинного мозга и позвонков. Краниально твердая оболочка срастается с краями большого отверстия затылочной кости, а каудально заканчивается на уровне II–III крестцовых позвонков, суживаясь в виде нити, f?lum d?rae m?tris spin?lis, которая прикрепляется к копчику.
Артериитвердая оболочка получает из спинномозговых ветвей сегментарных артерий,вены ее вливаются в pl?xus ven?sus vertebr?lis int?rnus, а нервы ее происходят из r?mi men?ngei спинномозговых нервов. Внутренняя поверхность твердой оболочки покрыта слоем эндотелия, вследствие чего имеет гладкий блестящий вид.
2. Паутинная оболочка спинного мозга, arachno?dea spin?lis, в виде тонкого прозрачного бессосудистого листка прилегает изнутри к твердой оболочке, отделяясь от последней щелевидным, пронизанным тонкими перекладинами субдуральным пространством, sp?tium subdur?le. Между паутинной оболочкой и непосредственно покрывающей спинной мозг мягкой оболочкой находится подпаутинное пространство, c?vitas subarachnoid?lis, в котором мозг и нервные корешки лежат свободно, окруженные большим количеством спинномозговой жидкости, liquor сегеbrospin?lis. Это пространство в особенности широко в нижней части арахноидального мешка, где оно окружает c?uda equ?na спинного мозга (cist?rna termin?lis). Наполняющая подпаутинное пространство жидкость находится в непрерывном сообщении с жидкостью подпаутинных пространств головного мозга и мозговых желудочков. Между паутинной оболочкой и покрывающей спинной мозг мягкой оболочкой в шейной области сзади, вдоль средней линии образуется перегородка, s?ptum cervic?le interm?dium. Кроме того, по бокам спинного мозга во фронтальной плоскости располагается зубчатая связка, lig. denticul?tum, состоящая из 19–23 зубцов, проходящих в промежутках между передними и задними корешками. Зубчатые связки служат для укрепления мозга на месте, не позволяя ему вытягиваться в длину. Посредством обеих ligg. denticul?tae подпаутинное пространство делится на передний и задний отделы.
3. Мягкая оболочка спинного мозга, p?a m?ter spin?lis, покрытая с поверхности эндотелием, непосредственно облекает спинной мозг и содержит между двумя своими листками сосуды, вместе с которыми заходит в его борозды и мозговое вещество, образуя вокруг сосудов периваскулярные лимфатические пространства.
Сосуды спинного мозга (рис. 271). Аа. spin?les ant?rior et post?rior, спускаясь вдоль спинного мозга, соединяются между собой многочисленными ветвями, образуя на поверхности мозга сосудистую сеть (так называемую vasocor?na). От этой сети отходят веточки, проникающие вместе с отростками мягкой оболочки в вещество мозга (см. рис. 271). Вены в общем аналогичны артериям и впадают в конечном итоге в pl?xus ven?si vertebr?les int?mi. К лимфатическим сосудам спинного мозга можно отнести периваскулярные пространства вокруг сосудов, сообщающиеся с подпаутинным пространством.
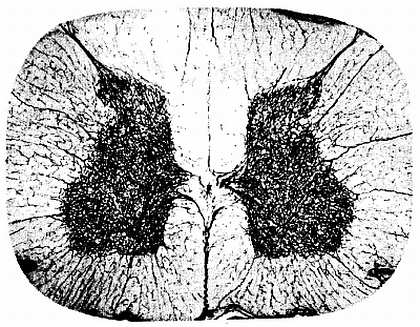
Рис. 271
ГОЛОВНОЙ МОЗГ
ОБЩИЙ ОБЗОР ГОЛОВНОГО МОЗГА
Головной мозг, enc?phalon, помещается в полости черепа и имеет форму, в общих чертах соответствующую внутренним очертаниям черепной полости. Его верхнелатеральная, или дорсальная, поверхность сообразно своду черепа выпукла, а нижняя, или основание мозга, более или менее уплощена и неровна. В головном мозге можно различить три крупные части: большой мозг (c?rebrum), мозжечок (cereb?llum) и мозговой ствол (tr?ncus enceph?licus). Наибольшую часть всего головного мозга занимают полушария большого мозга, за ними по величине следует мозжечок, остальную, сравнительно небольшую, часть составляет мозговой ствол.
Верхнелатеральная поверхность полушарий большого мозга. Оба полушария отделяются друг от друга щелью, fiss?ra longitudin?lis c?rebri, идущей в сагиттальном направлении. В глубине продольной щели полушария связаны между собой спайкой — мозолистым телом, c?rpus call?sum, и другими лежащими под ним образованиями. Спереди от мозолистого тела продольная щель сквозная, а сзади она переходит в поперечную щель мозга fiss?ra transv?rsa c?rebri, отделяющую задние части полушарий от лежащего под ними мозжечка.
Нижняя поверхность полушарий большого мозга (рис. 272).
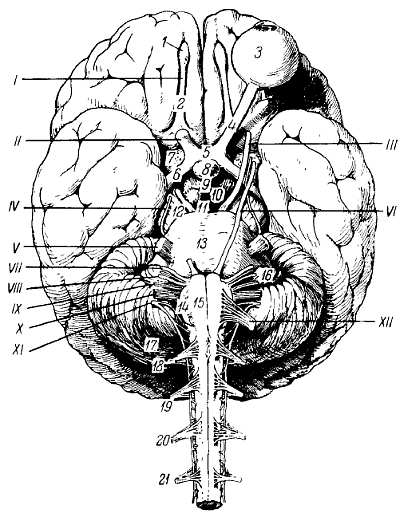
Рис. 272.
I — tr. olfactorius; II — n. opticus; III — n. oculomotorius; IV — n. trochlearis; V — n. trigeminus; VI — n. abducens; VII— n. facialis; VIII — n. vestibulocochlearis; IX — n. glossopharyngeus; X — n. vagus; XI — n. accessorius; XII — n. hypoglossus; 1 — bulbus olfactorius; 2 — tr. olfactorius; 3 — bulbus oculi sinistri; 4 — n. opticus; 5 — chiasma opticum; 6 — tr. opticus; 7 — substantia perforata anterior; 8 — hypophysis; 9 — tuber cinereum; 10 — corpus mamillare; 11 — substantia perforata posterior; 12 — pedunculus cerebri; 13 — pons; 14 — oliva; 15 — pyramis; 16 — flocculus; 17 — cerebellum; 18, 19, 20, 21 — корешки четырех верхних спинномозговых нервов.
Со стороны нижней поверхности мозга, f?cies inf?rior c?rebri, видна не только нижняя сторона полушарий большого мозга и мозжечка, но и вся нижняя поверхность мозгового ствола, а также отходящие от мозга нервы.
Передний отдел нижней поверхности головного мозга представлен лобными долями полушарий. На нижней поверхности лобных долей замечаютсяобонятельные луковицы, b?lbi olfact?rii, к которым из полости носа через отверстия l?mina cribr?sa решетчатой кости подходят тонкие нервные нити, f?lа оIfact?ria, образующие в своей совокупности I пару черепных нервов — обонятельные нервы, nn. olfact?rii. Обыкновенно при вынимании мозга из черепа эти нити отрываются от b?lbus olfact?rius.
Обонятельные луковицы продолжаются кзади в обонятельные тракты, tr?ctus olfact?rii, оканчивающиеся каждый двумя корешками, между которыми находится возвышение, называемое trig?num olfact?rium. Непосредственно сзади последнего на той и другой стороне находитсяпереднее продырявленное вещество, subst?ntia perfor?ta ant?rior, названное так по причине наличия здесь маленьких дырочек, через которые проходят в мозговое вещество сосуды.
Посередине между обоими передними продырявленными пространствами лежит зрительный перекрест, chi?sma ?pticum, имеющий форму буквы «X». От верхней поверхности хиазмы отходит тоненькая пластинка серого цвета, l?mina termin?lis, идущая в глубь fiss?ra longitudin?lis c?rebri. Сзади зрительного перекрестка помещаетсясерый бугор, t?bег ciner?um; верхушка его вытянута в узкую трубку, так называемую воронку, infundibulum, к которой подвешен расположенный в турецком седле гипофиз, hyp?physis c?rebri. Позади серого бугра находятся два шарообразных, белого цвета возвышения — сосцевидные тела, c?rpora mamill?ria. За ними лежит довольно глубокая межножковая ямка, f?ssa interpeduncul?ris, ограниченная с боков двумя толстыми валиками, сходящимися кзади и называемыми ножками мозга, ped?nculi c?rebri. Дно ямки пронизано отверстиями для сосудов, а потому носит название заднего продырявленного вещества, subst?ntia perfor?ta post?rior. Рядом с этим веществом в борозде медиального края мозговой ножки на той и другой стороне выходит III пара — глазодвигательный нерв, n. oculomot?ris. Сбоку ножек мозга виден самый тонкий из черепных нервов — блоковый нерв, n. trochle?ris — IV пара, который, однако, отходит не на основании мозга, а с его дорсальной стороны, из так называемого верхнего мозгового паруса. Позади ножек мозга находится толстый поперечный вал — мост, pons, который, суживаясь с боков, погружается в мозжечок. Боковые части моста, ближайшие к мозжечку, носят названиесредних ножек мозжечка, ped?nculi cerebell?res m?dii; на границе между ними и собственно мостом выхолит на той и другой стороне V пара — тройничный нерв, n. trig?minus. Позади моста лежит продолговатый мозг, med?la oblong?ta; между ним и задним краем моста по бокам средней линии видно начало VI пары —отводящего нерва, n. abd?cens; еще далее вбок у заднего края средних ножек мозжечка выходят рядом на той и другой стороне еще два нерва: VII — пара — лицевой нерв n. faci?lis, и VIII пара — n. vestibulocochle?ris.
Между пирамидой и оливой продолговатого мозга выходят корешки XII пары — подъязычного нерва, n. hypogl?ssus. Корешки IX, X и XI пар — n. glossophar?ngeus, n. v?gus и n. access?rius (верхняя часть) — выходят из бороздки позади оливы. Нижние волокна XI пары отходят уже от спинного мозга в шейной его части.
ЭМБРИОГЕНЕЗ ГОЛОВНОГО МОЗГА[34]
Нервная трубка очень рано подразделяется на два отдела, соответствующие головному и спинному мозгу. Передний, расширенный ее отдел, представляющий зачаток головного мозга, как отмечалось, расчленяется путем перетяжек на три первичных мозговых пузыря, лежащих друг за другом: передний, prosenc?phalon, средний, mesenc?phalon, и задний, rhombenc?phalon. Передний мозговой пузырь замыкается спереди так называемой концевой пластинкой, l?mina termin?lis. Эта стадия из трех пузырей при последующей дифференцировке переходит в стадию пяти пузырей, дающих начало пяти главным отделам головного мозга (рис. 273). Одновременно с этим мозговая трубка изгибается в сагиттальном направлении. Прежде всего в области среднего пузыря развивается выпуклый в дорсальную сторону головной изгиб, а затем на границе с зачатком спинного мозга также выпуклый дорсально шейный изгиб. Между ними образуется в области заднего пузыря третий изгиб, выпуклый в вентральную сторону, — мостовой изгиб.
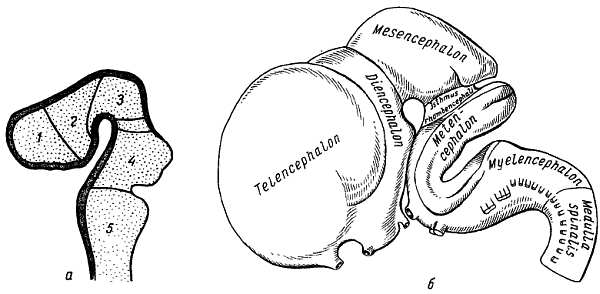
Рис. 273.
а — пять мозговых пузырей: 1 — telencephalon — конечный мозг; 2 — diencephalon — промежуточный мозг; 3 — mesencephalon — средний мозг; 4 — metencephalon — собственно задний мозг как часть ромбовидного мозга; 5 — myelencephalon — продолговатый мозг; между 3-м и 4-м пузырем — перешеек (isthmus rhombencephali); б — развитие головного мозга (по Р. Д. Синельникову).
Посредством этого последнего изгиба задний мозговой пузырь, rhombenc?phalon, делится на два отдела. Из них задний, myelenc?phalon, превращается при окончательном развитии в продолговатый мозг, а из переднего отдела, называемого metenc?phalon, развивается с вентральной стороны мост и с дорсальной стороны мозжечок. Metenc?phalon отделяется <рт лежащего впереди него пузырька среднего мозга узкой перетяжкой,?sthmus rhomben?ephali. Оощая полость rhombenc?phalon, имеющая на горизонтальном сечении вид ромба, образует IV желудочек, сообщающийся с центральным каналом спинного мозга. Вентральная и боковые стенки ее благодаря развитию в них ядер черепных нервов сильно утолщаются, дорсальная же стенка остается тонкой. В области продолговатого мозга большая часть ее состоит только из одного эпителиального слоя, срастающегося с мягкой оболочкой (t?la choro?dea inf?rior). Стенки среднего мозгового пузыря, mesenc?phalon, утолщаются при развитии в них мозгового вещества более равномерно. Вентрально из них возникают ножки мозга, а с дорсальной стороны — крыша среднего мозга (см. рис. 273). Полость среднего пузыря превращается в узкий канал — водопровод, соединяющийся с IV желудочком.
Более значительной дифференцировке и видоизменениям в форме подвергается передним мозговой пузырь, prosenc?phalon, который подразделяется на заднюю часть, dienc?phalon (промежуточный мозг), и переднюю, telenc?phalon (конечный мозг). Боковые стенки промежуточного мозга, утолщаясь, образуют таламусы (thal?mi). Кроме того, боковые стенки, выпячиваясь в стороны, образуют два зрительных пузырька, из которых впоследствии развиваются сетчатка глаз и зрительные нервы. Дорсальная стенка промежуточного мозга остается тонкой, в виде эпителиальной пластинки, срастающейся с мягкой оболочкой (t?la choro?dea sup?rior). Сзади из этой стенки возникает выпячивание, за счет которого происходит шишковидное тело (c?rpus pine?le). Полые ножки глазных пузырьков втягиваются с вентральной стороны в стенку переднего мозгового пузыря, вследствие чего на дне полости последнего образуется углубление, rec?ssus ?pticus, передняя стенка которого состоит из тонкой l?mina termin?lis. Позади rec?ssus ?pticus возникает другое воронкообразное углубление, стенки которого дают t?ber ciner?um, infund?bulum и заднюю (нервную) долю hyp?physis c?rebri. Еще далее кзади в области dienc?phalon в виде одиночного возвышения закладываются парные c?rpora mamill?ria. Полость промежуточного мозга образует III желудочек.
Telenc?phalon разделяется на срединную, меньшую, часть (pars mediana) и две большие боковые части — полушария большого мозга (hemisph?ria d?xtrum et sin?strum), которые у человека разрастаются очень сильно и в конце развития по величине значительно превосходят остальные отделы головного мозга. Полость pars medi?na, являющаяся передним продолжением полости промежуточного мозга (III желудочек), по бокам сообщается посредством межжелудочковых отверстий с полостями полушарий, которые на развитом мозге носят название боковых желудочков. Передняя стенка, представляющая непосредственное продолжение l?mina termin?lis, в начале первого месяца эмбриональной жизни образует утолщение, так называемую комиссуральную пластинку, из которой впоследствии развиваются мозолистое тело и передняя комиссура.
В основании каждого полушария, внутри, образуется выступ, так называемая полосатая часть, из которой развивается полосатое тело, c?rpus stri?tum. Часть медиальной стенки полушарий остается в виде одного эпителиального слоя, который вворачивается внутрь пузырька складкой мягкой оболочки (pl?xus choro?deus). На нижней стороне каждого полушария уже на 5-й неделе эмбриональной жизни образуется выпячивание — зачаток обонятельного мозга, rhinenс?phalon, которое постепенно отграничивается от стенки полушарий бороздкой, соответствующей fiss?ra rhin?lis later?lis. При развитии серого вещества (коры), а затем и белого в стенках полушария последнее увеличивается и образует так называемый плащ, p?llium, лежащий над обонятельным мозгом и покрывающий собой не только таламусы, но и дорсальную поверхность среднего мозга и мозжечка.
Полушарие при своем росте увеличивается сначала в области лобной доли, затем теменной и затылочной и, наконец, височной. Благодаря этому создается впечатление, как. будто плащ вращается вокруг таламусов сначала спереди назад, затем вниз и, наконец, загибается вперед, к лобной доле. Вследствие этого на боковой поверхности полушария, между лобной долей и приблизившейся к ней височной, образуется ямка, f?ssa later?lis c?rebri, которая при сближении названных долей большого мозга превращается в щель, s?lcus c?rebri later?lis; на дне ее образуется островок, ?nsula.
При развитии и росте полушария вместе с ним развиваются и совершают указанное «вращение» и его внутренние камеры, боковые желудочки мозга, а также часть c?rpus stri?tum (хвостатое ядро), чем и объясняется сходство их формы с формой полушария: у желудочков — наличие передней, центральной и задней частей и загибающейся книзу и вперед нижней части (см. рис. 295), у хвостатого ядра — наличие головки, тела и загибающегося книзу и вперед хвоста.
Борозды и извилины (рис. 274, 275, 276) возникают вследствие неравномерного роста самого мозга, что связано с развитием отдельных его частей.
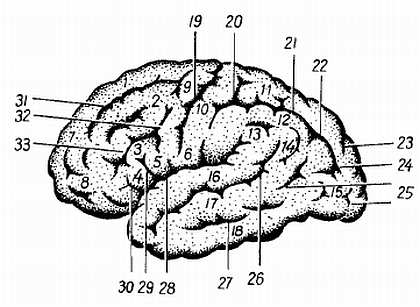
Рис. 274.
1, 7 — gyrus frontalis superior; 2 — gyrus frontalis medius; 3 — gyrus frontalis inferior; 4 — pars triangularis; 5 — pars opercularis; 8 — polus frontalis; 9 — gyrus precentralis; 10, 6 — gyrus postcentralis; 11 — lobulus parietalis superior; 12 — lobulus parietalis inferior; 13 — gyrus ssupramarginalis; 14 — gyrus angularis; 15 — gyri occipitales laterales; 16 — gyrus temporafis superior; 17 — gyrus temporalis medius; 18 — gyrus temporalis inferior; 19 — sul. centralis; 20 — sul. postcentralis; 21 — sul. intraparietalis; 22 — соединение sul. intraparietalis и sul. occipitalis transversus; 23 — sul. parietoccipitalis; 24 — sul. occipitalis transversus; 25 — sull. occipitales laterales; 26 — sul. temporalis superior; 27 — sul. temporalis inferior; 28 — sill. cerebri lateralis; 29 — r. ascendens sulci cerebri lateralis; 30 — r. anterior sulci cerebri lateralis; 31 — sul. frontalis superior; 32 — sul. precentralis; 33 — sul. frontalis inferior.
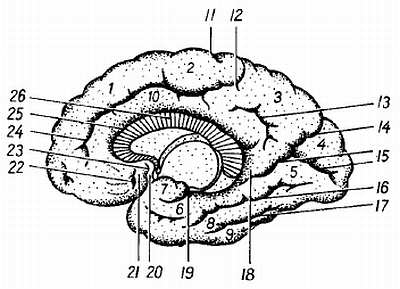
Рис. 275.
1 — gyrus frontalis superior; 2 — lobulus paracentralis; 3 — precuneus; 4 — cuneus; 5 — gyrus lingualis; 6 — gyrus parahippocampalis; 7 — uncus; 8 — gyrus occipitotemporalis medialis; 9 — gyrus occipitotemporalis lateralis; 10 — gyrus cinguli; 11 — sul. centralis; 12, 24 — sul. cinguli; 13 — sul. subparietalis; 14 — sul. parietooccipitalis; 15 — sul. calcarinus; 16 — sul. collateralis; 17 — sul. occipitotemporalis; 18 — isthmus gyri cinguli; 19 — sul. hippocampi; 20 — gyrus subcallosus; 21 — area olfactoria; 22 — sul. olfaetorius anterior; 23 — sul. olfactorius posterior; 25 — sul. corporis callosi; 26 — corpus callosum.
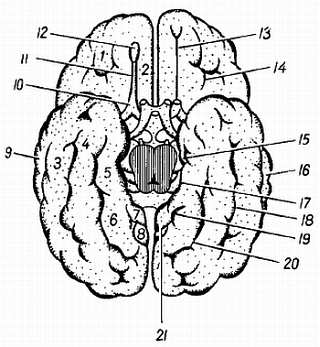
Рис. 276
1 — gyri orbitales; 2 — gyrus rectus; 3, 4 — gyri occipitotemporales medialis et lateralis; 5 — gyrus parahippocampalis; 6 — gyrus occipitotemporalis medialis; 7 — isthmus gyri cinguli; 8 — cuneus; 9 — gyrus temporalis medius; 10 — trigonum olfactorium; 11 — tr. olfactorius; 12 — bulbus olfactorius; 13 — sul. olfactorius; 14 — sulci orbitales; 15 — uncus gyri parahippocampalis; 16 — sul. temporalis inferior; 17 — sul. hippocampi; 18 — sul. occipitotemporalis; 19 — sul. calcarinus; 20 — sul. collateralis; 21 — sul. parietooccipitalis.
Так, на месте обонятельного мозга возникают s?lcus olfact?rius, s?lcus hippocampi и s?lcus c?nguli; на границе корковых концов кожного и двигательного анализаторов (понятие анализатора и описание борозд см. ниже) — s?lcus centr?lis; на границе двигательного анализатора и премоторной зоны, получающей импульсы от внутренностей, — s?lcus precentr?lis; на месте слухового анализатора — s?lcus tempor?lis sup?rior; в области зрительного анализатора — s?lcus calcarinus и s?lcus parietooccipit?lis.
Все эти борозды, появляющиеся раньше других и отличающиеся абсолютным постоянством, относятся к первичным бороздам. Остальные борозды, имеющие наименования и также возникающие в связи с развитием анализаторов, но появляющиеся несколько позднее и отличающиеся меньшим постоянством, относятся ко вторичным бороздам. К моменту рождения имеются все борозды — первичные и вторичные. Наконец, многочисленные мелкие бороздки, не имеющие названий, появляются не только в утробной жизни, но и после рождения. Они крайне непостоянны по времени появления, месту и числу; это третичные борозды. От степени их развития зависят все разнообразие и сложность мозгового рельефа. Рост человеческого мозга в эмбриональном периоде и в первые годы жизни, пока идут бурный рост организма, его приспособление к новой среде, приобретение способности к прямохождению и становление второй, словесной, сигнальной системы, происходит очень интенсивно и заканчивается к 20 годам. У новорожденных мозг (в среднем) массой 340 г у мальчиков и 330 г у девочек, а у взрослого — 1375 г у мужчин и 1245 г у женщин.
ОТДЕЛЬНЫЕ ЧАСТИ ГОЛОВНОГО МОЗГА
На основании эмбрионального развития, как было уже указано, головной мозг делится на отделы, располагающиеся, начиная с каудального конца, в таком порядке:
1) rhombenc?phalon — ромбовидный, или задний, мозг, который в свою очередь состоит из: a) myelenc?phalon — продолговатого мозга и б) mеlenc?phalon — собственно заднего мозга; 2) mesenc?phalon — средний мозг; 3) prosenc?phalon — передний мозг, в котором различают: a) dienc?phalon — промежуточный мозг и б) telenc?phalon — конечный мозг.
Все названные отделы, кроме мозжечка и конечного мозга, составляют мозговой ствол.
Кроме таких отделов, выделяют еще перешеек, isthmus rhombenc?phali, между rhombenc?phalon и средним мозгом.
Prosenc?phalon составляет большой мозг — c?rebrum в отличие от малого мозга — мозжечка, cereb?llum.
Ромбовидный мозг
Продолговатый мозг
Продолговатый мозг, myelenc?phalon, med?lla oblong?ta (рис. 277, 278), представляет непосредственное продолжение спинного мозга в ствол головного мозга и является частью ромбовидного мозга. Он сочетает в себе черты строения спинного мозга и начального отдела головного, чем и оправдывается его название myelenc?phalon. Med?lla oblong?ta имеет вид луковицы, b?lbus c?rebri (отсюда термин «бульбарные расстройства»); верхний расширенный конец граничит с мостом, а нижней границей служит место выхода корешков 1 пары шейных нервов или уровень большого отверстия затылочной кости.
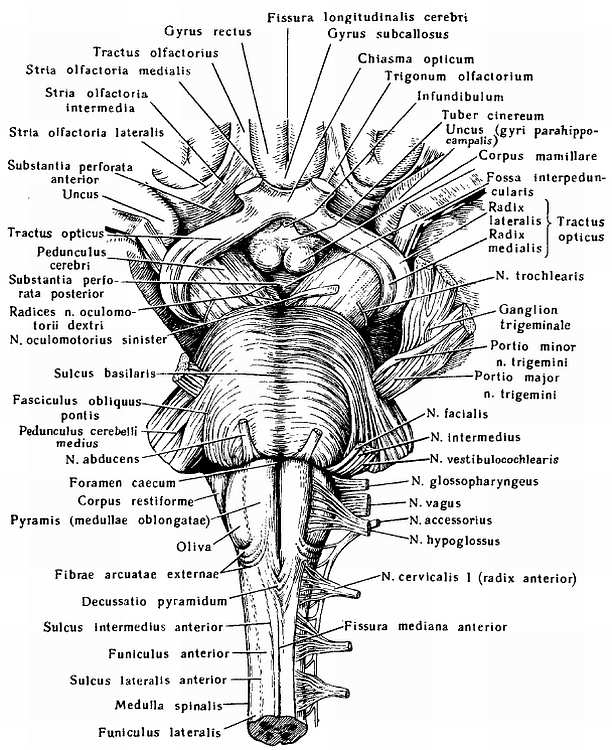
Рис. 277.
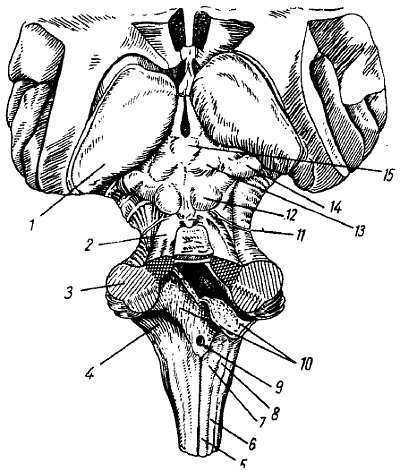
Рис. 278
1 — pulvinar (задняя часть thalamus); 2 — pedunculus cerebellaris superior; 3 — pedunculus cerebellaris medius; 4 — pedunculus cerebellaris inferior; 5 — fasc. gracilis; 6 — fasc. cuneatus; 7 — tuberculum gracilum; 8 — tuberculum cuneatum; 9 — apertura meaiana ventriculi quarti; 10 — plexus chorloideus и tela chorioidea ventriculi quarlti (разрезаны и отвернуты, через разрез видна полость IV желудочка); 11 — n. trochlearis; 12 — colliculus inferior крыши среднего мозга; 13 — colliculus superior крыши среднего мозга; 14 — corpus geniculatum mediate; 15 — corpus pineale.
1. На передней (вентральной) поверхности продолговатого мозга по средней линии проходит fiss?ra medi?na ant?rior, составляющая продолжение одноименной борозды спинного мозга. По бокам ее на той и другой стороне находятся два продольных тяжа — пирамиды, pyram?des med?llae oblong?tae, которые как бы продолжаются в передние канатики спинного мозга. Составляющие пирамиды пучки нервных волокон частью перекрещиваются в глубине fiss?ra medi?na ant?rior с аналогичными волокнами противоположной стороны — decuss?tio pyram?dum, после чего спускаются в боковом канатике на другой стороне спинного мозга — tr?ctus corticospin?lis (pyramid?lis) later?lis, частью остаются неперекрещенными и спускаются в переднем канатике спинного мозга на своей стороне — tr?ctus corticospin?lis (pyramid?lis) ant?rior.
Пирамиды отсутствуют у низших позвоночных и появляются по мере развития новой коры; поэтому они наиболее развиты у человека, так как пирамидные волокна соединяют кору большого мозга, достигшую у человека наивысшего развития, с ядрами черепных нервов и передними рогами спинного мозга.
Латерально от пирамиды лежит овальное возвышение — оливa, ol?va, которая отделена от пирамиды бороздкой, s?lcus anterolater?lis.
2. На задней (дорсальной) поверхности продолговатого мозга (см. рис. 278) тянется s?lcus medi?nus post?rior — непосредственное продолжение одноименной борозды спинного мозга. По бокам ее лежат задние канатики, ограниченные латерально с той и другой стороны слабо выраженной s?lcus posterolater?lis. По направлению кверху задние канатики расходятся в стороны и идут к мозжечку, входя в состав его нижних ножек, ped?nculi cerebell?res inferi?res, окаймляющих снизу ромбовидную ямку. Каждый задний канатик подразделяется при помощи промежуточной борозды на медиальный, fasc?culus gr?cilis, и латеральный, fasc?culus cune?tus. У нижнего угла ромбовидной ямки тонкий и клиновидный пучки приобретают утолщения — tub?rculum gr?cilum и tub?rculum cune?tum. Эти утолщения обусловлены соименными с пучками ядрами серого вещества, n?cleus gr?cilis и n?cleus cune?tus. В названных ядрах оканчиваются проходящие в задних канатиках восходящие волокна спинного мозга (тонкий и клиновидный пучки). Латеральная поверхность продолговатого мозга, находящаяся между s?lci posterolater?lis et anterolater?lis, соответствует боковому канатику. Из s?lcus posterolater?lis позади оливы выходят XI, X и IX пары черепных нервов. В состав продолговатого мозга входит нижняя часть ромбовидной ямки (рис. 279; см. рис. 283).
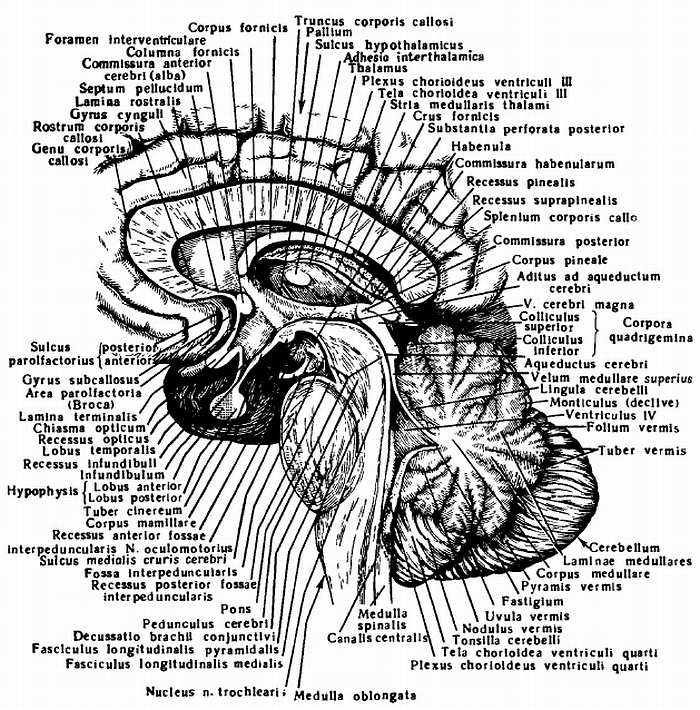
Рис. 279.
Внутреннее строение продолговатого мозга. Продолговатый мозг возник в связи с развитием органов гравитации и слуха, а также в связи с жаберным аппаратом, имеющим отношение к дыханию и кровообращению. Поэтому в нем заложены ядра серого вещества, имеющие отношение к равновесию, координации движений, а также к регуляции обмена веществ, дыхания и кровообращения (рис. 280).
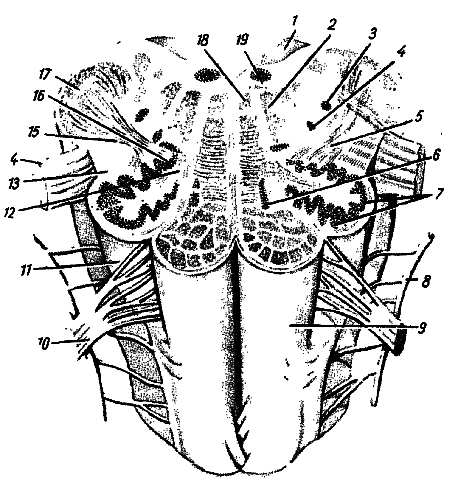
Рис. 280.
1 — velum medullare posterior; 2 — formatio reticularis; 3 — nucl. tracius spinalis n. trigemini; 4 — nucl. ambiguus; 5 — tr. olivocerebellaris; 6 — nucl. olivaris accessorius medialis; 7, 16 — nucll. olivares; 8 — n. accessorius; 9 — pyramis; 10 — n. hypoglossus; 11 — oliva; 12 — hilus nuclei olivaris; 13 — tr. tectospinalis; 14 — n. vagus; 15 — tr. rubrospinalis; 17 — pedunculus cerebellaris inferior; 18 — fasc. longitudinalis medialis; 19 — nucl. n. hypoglossi.
1. N?cleus oliv?ris, ядро оливы, имеет вид извитой пластинки серого вещества, открытой медиально (h?lus), и обусловливает снаружи выпячивание оливы. Оно связано с зубчатым ядром мозжечка и является промежуточным ядром равновесия, наиболее выраженным у человека, вертикальное положение которого нуждается в совершенном аппарате гравитации. (Встречается еще n?cleus oliv?ris access?rius medi?lis.)
2. Form?tio reticul?ris, ретикулярная формация, образующаяся из переплетения нервных волокон и лежащих между ними нервных клеток.
3. Ядра четырех пар нижних черепных нервов (XII–IX), имеющие отношение к иннервации производных жаберного аппарата и внутренностей.
4. Жизненно важные центры дыхания и кровообращения, связанные с ядрами блуждающего нерва. Поэтому при повреждении продолговатого мозга может наступить смерть.
Белое вещество продолговатого мозга содержит длинные и короткие волокна. К длинным относятся проходящие транзитно в передние канатики спинного мозга нисходящие пирамидные пути, частью перекрещивающиеся в области пирамид. Кроме того, в ядрах задних канатиков (n?clei gr?cilis et cune?tus) находятся тела вторых нейронов восходящих чувствительных путей. Их отростки идут от продолговатого мозга к таламусу, tr?ctus bulbothal?micits. Волокна этого пучка образуют медиальную петлю, lemn?scus medi?lis, которая в продолговатом мозге совершает перекрест, decuss?tio lemnisc?rum, и в виде пучка волокон, расположенных дорсальнее пирамид, между оливами — межоливный петлевой слой — идет далее. Таким образом, в продолговатом мозге имеется два перекрестка длинных проводящих путей: вентральный двигательный, decuss?tio pyramidum, и дорсальный чувствительный, decuss?tio lemnisc?rum.
К коротким путям относятся пучки нервных волокон, соединяющие между собой отдельные ядра серого вещества, а также ядра продолговатого мозга с соседними отделами головного мозга. Среди них следует отметить tr?ctus olivocerebell?ris и лежащий дорсально от межоливного слоя fasc?culus longitudin?lis medi?lis.
Топографические взаимоотношения главнейших образований продолговатого мозга видны на поперечном срезе, проведенном на уровне олив (см. рис. 280). Отходящие от ядер подъязычного и блуждающего нервов корешки делят продолговатый мозг на той и другой стороне на три области: заднюю, боковую и переднюю. В задней лежат ядра заднего канатика и нижние ножки мозжечка, в боковой — ядро оливы и form?tio reticul?ris и в передней — пирамиды.
Задний мозг
Metencephalon состоит из двух частей: вентральной — моста и дорсальной — мозжечка.
Мост
Мост, pons, представляет собой со стороны основания мозга толстый белый вал, граничащий сзади с верхним концом продолговатого мозга, а спереди — с ножками мозга (см. рис. 277). Латеральной границей моста служит искусственно проводимая линия через корешки тройничного и лицевого нервов, linea trigeminofaci?lis. Латерально от этой линии находятся средние мозжечковые ножки, ped?nculi cerebell?res m?dii, погружающиеся на той и другой стороне в мозжечок. Дорсальная поверхность моста не видна снаружи, так как она скрыта под мозжечком, образуя верхнюю часть ромбовидной ямки (дна IV желудочка). Вентральная поверхность моста имеет волокнистый характер, причем волокна в общем идут поперечно и направляются в ped?nculi cerebell?res m?dii. По средней линии вентральной поверхности проходит пологая канавка, s?lcus basil?ris, в которой лежит a. basil?ris.
Внутреннее строение моста. На поперечных разрезах моста можно видеть, что он состоит из большей передней, или вентральной, части, pars ventr?lis p?ntis, и меньшейдорсальной, pars dors?lis p?ntis. Границей между ними служит толстый слой поперечных волокон — трапециевидное тело, c?rpus trapezo?deum, волокна которого относятся к слуховому пути. В области трапециевидного тела располагается ядро, также имеющее отношение к слуховому пути, — n?cleus dors?lis c?rporis trapezo?dei.
Pars ventr?lis содержит продольные и поперечные волокна, между которыми разбросаны собственные ядра серого вещества, n?clei p?ntis.
Продольные волокна принадлежат к пирамидным путям, к fibrae corticopontinae, которые связаны с собственными ядрами моста, откуда берут начало поперечные волокна, идущие к коре мозжечка, tr?ctus pontocerebell?ris. Вся эта система проводящих путей связывает через мост кору полушарий большого мозга с корой полушарий мозжечка. Чем сильнее развита кора большого мозга, тем сильнее развиты мост и мозжечок. Естественно, что мост оказывается наиболее выраженным у человека, что является специфический чертой строения его головного мозга. В pars dors?lis находится form?tio reticul?ris p?ntis, являющееся продолжением такой же формации продолговатого мозга, а поверх ретикулярной формации — выстланное эпендимой дно ромбовидной ямки с лежащими под ним ядрами черепных нервов (VIII–V пары).
В pars dors?lis продолжаются также проводящие пути продолговатого мозга, располагающиеся между средней линией и n?cleus dors?lis c?rporis trapezo?dei и входящие в состав медиальной петли, lemn?scus medi?lis; в последней перекрещиваются восходящие пути продолговатого мозга, tr?ctus bulbothal?micus.
Мозжечок
Мозжечок, cereb?llum, является производным заднего мозга, развившегося в связи с рецепторами гравитации. Поэтому он имеет прямое отношение к координации движений и является органом приспособления организма к преодолению основных свойств массы тела — тяжести и инерции.
Развитие мозжечка в процессе филогенеза прошло 3 основных этапа соответственно изменению способов передвижения животного.
Мозжечок впервые появляется в классе круглоротых, у миног, в виде поперечной пластинки. У низших позвоночных (рыбы) выделяются парные ушковидные части (archicerebellum) и непарное тело (paleocerebellum), соответствующее червю; у пресмыкающихся и птиц сильно развито тело, а ушковидные части превращаются в рудиментарные. Полушария мозжечка возникают только у млекопитающих (neocerebellum). У человека в связи с прямохождением при помощи одной пары конечностей (ног) и усовершенствованием хватательных движений руки при трудовых процессах полушария мозжечка достигают наибольшего развития, так что мозжечок у человека развит сильнее, чем у всех животных, что составляет специфическую человеческую черту его строения.
Мозжечок помещается под затылочными долями полушарий большого мозга, дорсально от моста и продолговатого мозга, и лежит в задней черепной ямке. В нем различают объемистые боковые части, илиполушария, hemisph?ria cereb?lli, и расположенную между ними среднюю узкую часть — червь, v?rmis.
На переднем краю мозжечка находится передняя вырезка, которая охватывает прилежащую часть ствола мозга. На заднем краю имеется более узкая задняя вырезка, отделяющая полушария друг от друга.
Поверхность мозжечка покрыта слоем серого вещества, составляющим кору мозжечка, и образует узкие извилины — листки мозжечка, f?lia cereb?lli, отделенные друг от друга бороздами, fiss?rae cereb?lli. Среди них самая глубокая fiss?ra horizont?lis cereb?lli проходит по заднему краю мозжечка, отделяет верхнюю поверхность полушарий, f?cies sup?rior, от нижней, f?cies inf?rior. С помощью горизонтальной и других крупных борозд вся поверхность мозжечка делится на ряд долек, l?buli cereb?lli. Среди них необходимо выделить наиболее изолированную маленькую дольку — клочок, fl?cculus, лежащую на нижней поверхности каждого полушария у средней мозжечковой ножки, а также связанную с клочком часть червя — n?dulus, узелок. Fl?cculus соединен с n?dulus посредством тонкой полоски — ножки клочка, ped?nculus fl?cculi, которая медиально переходит в тонкую полулунную пластинку — нижний мозговой парус, v?lum medull?re inf?rius.
Внутреннее строение мозжечка. В толще мозжечка имеются парные ядра серого вещества, заложенные в каждой половине мозжечка среди белого ее вещества (рис. 281). По бокам от средней линии в области, где в мозжечок вдается шатер, fast?gium, лежит самое медиальное ядро — ядро шатра, n?cleus fast?gii. Латеральнее от него расположено шаровидное ядро, n?cleus glob?sus, а еще латеральнее — пробковидное ядро, n?cleus embolif?rmis. Наконец, в центре полушария находится зубчатое ядро, n?cleus dent?tus, имеющее вид серой извилистой пластинки, похожей на ядро оливы. Сходство n?cleus dent?tus мозжечка с имеющим также зубчатую форму ядром оливы не случайно, так как оба ядра связаны проводящими путями, f?brae olivocerebell?res, и каждая извилина одного ядра аналогична извилине другого. Таким образом, оба ядра вместе участвуют в осуществлении функции равновесия (см. рис. 280, 281).

Рис. 281.
1 — nucl. fastigii; 2 — nucl. globosus; 3 — nucl. emboliformis; 4 — nucl. dentatus.
Названные ядра мозжечка имеют различный филогенетический возраст: n?cleus fastigii относился к самой древней части мозжечка — fl?cculus (агchicereb?llum), связанной с вестибулярным аппаратом; n?clei embolif?rmis et glob?sus — к старой части (paleocereb?llum), возникшей в связи с движениями туловища, и n?cleus dent?tus — к самой молодой (neocereb?llum), развившейся в связи с передвижением при помощи конечностей. Поэтому при поражении каждой из этих частей нарушаются различные стороны двигательной функции, соответствующие различным стадиям филогенеза, а именно: при повреждении флоккулонодулярной системы и ее ядра шатра нарушается равновесие тела. При поражении червя и соответствующих ему пробковидного и шаровидного ядер нарушается работа мускулатуры шеи и туловища, при поражении полушарий и зубчатого ядра — работа мускулатуры конечностей.
Белое вещество мозжечка на разрезе имеет вид мелких листочков растения, соответствующих каждой извилине, покрытой с периферии корой серого вещества. В результате общая картина белого и серого вещества на разрезе мозжечка напоминает дерево, ?rbor vitae cereb?lli (древо жизни; название дано по внешнему виду, поскольку повреждение мозжечка не является непосредственной угрозой жизни). Белое вещество мозжечка слагается из различного рода нервных волокон. Одни из них связывают извилины и дольки, другие идут от коры к внутренним ядрам мозжечка и, наконец, третьи связывают мозжечок с соседними отделами мозга. Эти последние волокна идут в составе трех пар мозжечковых ножек:
1. Нижние ножки, ped?nculi cerebell?res inf?riores (к продолговатому мозгу). В их составе идут к мозжечку tr?ctus spinocerebell?ris post?rior, f?brae arcu?tae ext?rnae — от ядер задних канатиков продолговатого мозга и f?brae olivocerebell?res — от оливы. Первые два тракта оканчиваются в коре червя и полушарий. Кроме того, здесь идут волокна от ядер вестибулярного нерва, заканчивающиеся в nucleus fastigii. Благодаря всем этим волокнам мозжечок получает импульсы от вестибулярного аппарата и проприоцептивного поля, вследствие чего становится ядром проприоцеитивной чувствительности, совершающим автоматическую поправку на двигательную деятельность остальных отделов мозга. В составе нижних ножек идут также нисходящие пути в обратном направлении, а именно: от n?cleus fast?gii к латеральному вестибулярному ядру (см. ниже), а от него — к передним рогам спинного мозга, tr?ctus vestibulospin?lis. При посредстве этого пути мозжечок оказывает влияние на спинной мозг.
2. Средние ножки, pedunculi cerebellares medii (к мосту). В их составе идут нервные волокна от ядер моста к коре мозжечка. Возникающие в ядрах моста проводящие пути к коре мозжечка, tr?ctus pontocerebellares, находятся на продолжении корково-мостовых путей, f?brae corticopont?nae, оканчивающихся в ядрах моста после перекреста. Эти пути связывают кору большого мозга с корой мозжечка, чем и объясняется тот факт, что чем более развита кора большого мозга, тем более развиты мост и полушария мозжечка, что наблюдается у человека.
3. Верхние ножки, ped?nculi cerebell?res superi?res (к крыше среднего мозга). Они состоят из нервных волокон, идущих в обоих направлениях: 1) к мозжечку — tr?ctus spinocerebell?ris ant?rior и 2) от n?cleus dent?tus мозжечка к покрышке среднего мозга — tr?ctus cerebellotegment?lis, который после перекреста заканчивается в красном ядре и в таламусе. По первым путям в мозжечок идут импульсы от спинного мозга, а по вторым он посылает импульсы в экстрапирамидную систему, через которую сам влияет на спинной мозг.
Перешеек
Перешеек, ?sthmus rhombenc?phali, представляет переход от rhombenc?phalon к mesenc?phalon. В его состав входят: 1) верхние мозжечковые ножки, ped?nculi cerebell?res superi?res; 2) натянутый между ними и мозжечком верхний мозговой парус, v?lum medull?re sup?rius, который прикрепляется к срединной бороздке между холмиками пластинки крыши среднего мозга; 3) треугольник петли, trig?num lemn?sсi, обусловленный ходом слуховых волокон латеральной петли, lemn?scus later?lis. Этот треугольник серого цвета, ограничен спереди ручкой нижнего холмика, сзади — верхней ножкой мозжечка и латерально — ножкой мозга. Последняя отделена от перешейка и среднего мозга ясно выраженной бороздой, s?lcus later?lis mesenc?phali. Внутрь перешейка вдается верхний конец IV желудочка, переходящий в среднем мозге в водопровод.
IV желудочек
IV желудочек, ventriculus qu?rtus, представляет собой остаток полости заднего мозгового пузыря и поэтому является общей полостью для всех отделов заднего мозга, составляющих ромбовидный мозг, rhombenc?phalon (продолговатый мозг, мозжечок, мост и перешеек). IV желудочек напоминает палатку, в которой различают дно и крышу (см. рис. 279; рис. 282).
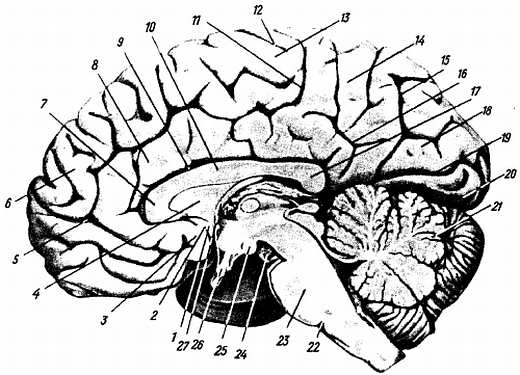
Рис. 282.
1 — gyrus subcallosus; 2 — sul. olfactorius posterior; 3 — area olfactoria; 4 — septum pellucidum; 5, 11 — sul. cinguli; 6 — gyrus frontalis superior; 7 — genu corporis callosi; 8 — gyrus einguli; 9 — sul. corporis callosi, 10 — tr. uncus corporis callosi; 12 — sul. centralis; 13 — lobulus paracentralis; 14 — precuneus; 15 — sul. parietooccipitalis; 16 — sul. subparietalis; 17 — splenium corporis callosi; 18 — cuneus; 19 — sul calcannus; 20 — gyrus occipitotemporalis medialis; 21 — долька нижнего червя; 22 — задняя граница моста; 23 — pons; 24 — n. oculomotonus; 25 — corpus mamillare; 26 — chiasma opticum; 27 — fornix (columna).
Дно, или основание, желудочка имеет форму ромба, как бы вдавленного в заднюю поверхность продолговатого мозга и моста. Поэтому его называют ромбовидной ямкой, f?ssa rhombo?dea. В задненижний угол ромбовидной ямки открывается центральный канал спинного мозга, а в передневерхнем углу IV желудочек сообщается с водопроводом. Латеральные углы заканчиваются слепо в виде двух карманов, rec?ssus later?les ventriculi qu?rti, загибающихся вентрально вокруг нижних ножек мозжечка.
Крыша IV желудочка, t?gmen ventr?culi qu?rti, имеет форму шатра и составлена двумя мозговыми парусами: верхним, v?lum medull?re sup?rius, натянутым между верхними ножками мозжечка, и нижним, v?lum medull?re inf?rius, парным образованием, примыкающим к ножкам клочка. Часть крыши между парусами образована веществом мозжечка. Нижний мозговой парус дополняется листком мягкой оболочки, t?la choro?dea ventr?culi qu?rti, покрытой изнутри слоем эпителия, l?mina choro?dea epitheli?lis, представляющим рудимент задней стенки заднего мозгового пузыря (с ней связано сплетение — pl?xus choro?deus ventr?culi qu?rti).
T?la choro?dea первоначально вполне замыкает полость желудочка, но затем в процессе развития в ней появляются три отверстия: одно в области нижнего угла ромбовидной ямки, apert?ra medi?na ventr?culi qu?rti (самое большое), и два в области боковых карманов желудочка, apert?rae later?les ventr?culi qu?rti. При посредстве этих отверстий IV желудочек сообщается с подпаутинным пространством головного мозга, благодаря чему спинномозговая жидкость поступает из мозговых желудочков в межоболочечные пространства. В случае сужения или заращения этих отверстий на почве воспаления мозговых оболочек (менингит) накапливающаяся в мозговых желудочках спинномозговая жидкость не находит себе выхода в подпаутинное пространство и возникает водянка головного мозга.
Ромбовидная ямка, f?ssa rhombo?dea (рис. 283), имеет соответственно ромбовидной форме четыре стороны — две верхние и две нижние.
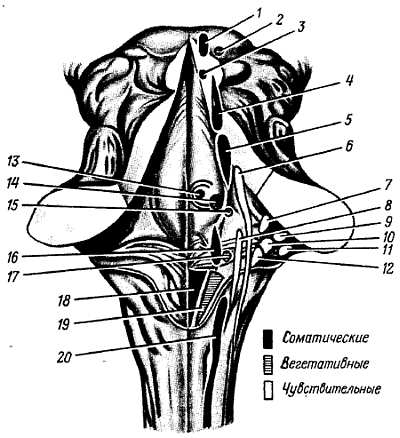
Рис. 283.
1 — nucl. n. oculomotorii; 2 — nucl. accessorius n. oculomotorii; 3 — nucl. n. trochlearis; 4 — nucl. mesencephalicus n. trigemini; 5 — nucl. motorius n. trigemini; 6 — nucl. pontis n. trigemini; 7 — nucl. vestibularis superior; 8 — nucl. n. solitarius (IX, X); 9 — nucl. cochlearis ventralis; 10 — nucl. vestibularis lateralis; 11 — nucl. cochlearis dorsalis; 12 — nucl. vestibularis medialis; 13 — nucl. abducentis; 14 — nucl. n. facialis; 15 — nucl. salivatorius superior; 16 — nucl. ambiguus; 17 — nucl. salivatorius inferior; 18 — nucl. n. hypoglossi; 19 — nucl. dorsalis n. vagi; 20 — nucl. n. iccessorii.
Верхние стороны ромба ограничены двумя верхними мозжечковыми ножками, а нижние стороны — двумя нижними ножками. Вдоль ромба, по средней линии, от верхнего угла к нижнему тянется срединная борозда, s?lcus medi?nus, которая делит ромбовидную ямку на правую и левую половины. По сторонам борозды расположено парное возвышение, emin?ntia medi?lis, обусловленное скоплением серого вещества.
Книзу emin?ntia medi?lis постепенно суживается, переходя в треугольник, на который проецируется ядро подъязычного нерва, trig?num n?rvi hypogl?ssi. Латеральнее нижней части этого треугольника лежит меньший треугольник, заметный по своей серой окраске, trig?num n?rvi v?gi, в котором заложено вегетативное ядро блуждающего нерва, n?cleus dors?lis n?rvi v?gi. Вверху eminentia medialis имеет возвышение — лицевой бугорок, coll?culus faci?lis, обусловленный прохождением корешка лицевого и проекцией ядра отводящего нервов.
В области латеральных углов располагается с обеих сторон вестибулярное поле, ?rea vestibul?ris, здесь помещаются ядра VIII пары. Часть выходящих из них волокон идет поперек ромбовидной ямки от латеральных углов к срединной борозде в виде горизонтальных полосок, str?ae medull?res ventr?culi qu?rti. Эти полоски делят ромбовидную ямку на верхнюю и нижнюю половины и соответствуют границе между продолговатым мозгом и мостом.
Топография серого вещества ромбовидной ямки. Серое вещество спинного мозга непосредственно переходит в серое вещество мозгового ствола и частью расстилается по ромбовидной ямке и стенкам водопровода (см. «Средний мозг»), а частью разбивается на отдельные ядра черепных нервов или ядра пучков проводящих путей.
Чтобы понять расположение этих ядер, нужно учитывать, как отмечалось, что замкнутая нервная трубка при переходе от спинного в продолговатый мозг раскрылась на своей задней стороне и развернулась в ромбовидную ямку. Вследствие этого задние рога серого вещества спинного мозга как бы разошлись в стороны. Заложенные в задних рогах соматически-чувствительные ядра расположились в ромбовидной ямке латерально, а соответствующие передним рогам соматически-двигательные ядра остались лежать медиально. Что касается вегетативных ядер, заложенных в боковых рогах спинного мозга, то соответственно положению боковых рогов между задними и передними эти ядра при развертывании нервной трубки оказались лежащими в ромбовидной ямке между соматически-чувствительными и соматически-двигательными ядрами. В результате в области ромбовидной ямки в отличие от спинного мозга ядра серого вещества расположены не в переднезаднем направлении, а лежат рядами — медиально и латерально.
Так, например, соматически-двигательные ядра XII и VI пар лежат в медиальном ряду, вегетативные ядра X, IX, VII пар — в среднем ряду и соматически-чувствительные ядра VIII пары — латерально.
Проекция ядер черепных нервов на ромбовидную ямку:
XII пара — подъязычный нерв, n. hypogl?ssus, имеет единственное двигательное ядро, заложенное в самой нижней части ромбовидной ямки, в глубине trig?num n. hypogl?ssi.
XI пара — добавочный нерв, n. access?rius, имеет два ядра (оба двигательные): одно заложено в спинном мозге и называется n?cleus n. асcess?rii, другое является каудальным продолжением ядер X и IX пар нервов и называется n?cleus ambiguus. Оно лежит в продолговатом мозге дорсолатералыю от ядра оливы.
X пара — блуждающий нерв, n. v?gus, имеет три ядра:
1) чувствительное ядро, n?cleus solit?rius, расположено рядом с ядром подъязычного нерва, в глубине trig?num n. v?gi;
2) вегетативное ядро, n?cleus dors?lis n. vagi, лежит в той же области;
3) двигательное ядро, n?cleus ambiguus (двойное), общее с ядром IX пары, заложено в form?tio reticul?ris, глубже n?cleus dors?lis.
IX пара — языкоглоточный нерв, n. glossophar?ngeus, также содержит три ядра:
1) чувствительное ядро, n?cleus solit?rius, лежит латеральнее ядра подъязычного нерва;
2) вегетативное (секреторное) ядро, n?cleus salivat?rius inf?rior, нижнее слюноотделительное ядро; клетки его рассеяны в form?tio reticul?ris продолговатого мозга между n. amb?guus и ядром оливы;
3) двигательное ядро, общее с n.v?gus и n.access?rius, n?cleus amb?guus.
VIII пара — преддверно-улитковый нерв, n. vestibulocochle?ris, имеет множественные ядра, проецирующиеся на латеральные углы ромбовидной ямки, в области ?rea vestibul?ris. Ядра делятся на две группы соответственно двум частям нерва. Одна часть нерва, pars cochle?ris, — нерв улитки, или собственно слуховой нерв, имеет два ядра: заднее, n?cleus cochle?ris dors?lis, и переднее, n?cleus cochle?ris ventr?lis, расположенное латеральнее и кпереди от предыдущего. Другая часть нерва, pars vestibul?ris, — нерв преддверия, или гравитационный нерв, имеет четыре ядра (n?clei vestibul?res):
1) медиальное — главное;
2) латеральное;
3) верхнее;
4) нижнее.
Наличие у человека четырех ядер отражает ранние стадии филогенеза, когда у рыб имелось несколько отдельных воспринимающих гравитационных аппаратов.
VII пара — лицевой нерв, n. faci?lis, имеет одно двигательное ядро, расположенное в form?tio reticul?ris p?rtis dors?lis моста. Отходящие от него нервные волокна на своем пути в толще моста образуют петлю, выпячивающуюся на ромбовидной ямке в виде coll?culus faci?lis.
Промежуточный нерв, n. interm?dius, тесно связанный в своем ходе с лицевым нервом, имеет два ядра:
1) вегетативное (секреторное), n?cleus salivat?rius sup?rior (верхнее слюноотделительное ядро), заложено в form?tio reticul?ris моста, дорсальнее ядра лицевого нерва;
2) чувствительное, n?cleus solit?rius.
VI пара — отводящий нерв, n. abd?cens. имеет одно двигательное ядро, заложенное в петле лицевого нерва, поэтому coll?culus faci?lis на поверхности ромбовидной ямки соответствует этому ядру.
V пара — тройничный нерв, n.trig?minus, имеет четыре ядра:
1) чувствительное, n?cleus pontinus n. trig?mini, проецируется в дорсолатеральной части верхнего отдела моста;
2) ядро спинномозгового тракта, n?cleus spin?lis n.trig?mini, является продолжением предыдущего по всему протяжению продолговатого мозга до шейного отдела спинного мозга, где соприкасается с subst?ntia gelatin?sa задних рогов;
3) двигательное ядро, n?cleus mot?rius n. trig?mini (жевательное), расположено медиальнее чувствительного;
4) ядро среднемозгового тракта, n?cleus mesenceph?licus n. trig?mini, лежит латеральнее водопровода. Оно представляет ядро проприоцептивной чувствительности для жевательных мышц и для мышц глазного яблока.
Возможно, что это ядро отражает самостоятельное развитие первой ветви тройничного нерва (n. ophth?lmicus), называемого у животных n. ophth?lmicus prof?ndus и имеющего отношение к органу зрения, чем и объясняется расположение ядра в среднем мозге.
Средний мозг
Средний мозг, mesenc?phalon, развивается в процессе филогенеза под преимущественным влиянием зрительного рецептора, поэтому важнейшие его образования имеют отношение к иннервации глаза. Здесь же образовались центры слуха, которые вместе с центрами зрения в дальнейшем разрослись в виде четырех холмиков крыши среднего мозга. С появлением у высших животных и человека коркового конца слухового и зрительного анализаторов в коре переднего мозга слуховые и зрительные центры среднего мозга сами попали в подчиненное положение и стали промежуточными, подкорковыми. С развитием у высших млекопитающих и человека переднего мозга через средний мозг стали проходить проводящие пути, связывающие кору конечного мозга со спинным (ножки мозга) (см. рис. 277, 278).
В результате в среднем мозге человека имеются: 1) подкорковые центры зрения и ядра нервов, иннервирующих мышцы глаза; 2) подкорковые слуховые центры; 3) все восходящие и нисходящие проводящие пути, связывающие кору головного мозга со спинным и идущие транзитно через средний мозг; 4) пучки белого вещества, связывающие средний мозг с другими отделами центральной нервной системы. Соответственно этому средний мозг, являющийся у человека наименьшим и наиболее просто устроенным отделом головного мозга, имеет две основные части: крышу, где располагаются подкорковые центры слуха и зрения, и ножки мозга, где преимущественно проходят проводящие пути.
1. Дорсальная часть, крыша среднего мозга, t?ctum mcsenc?phali.
Она скрыта под задним концом мозолистого тела и подразделяется посредством двух идущих крест-накрест канавок — продольной и поперечной — на четыре холмика, располагающихся попарно.
Верхние два холмика, coll?culi superi?res, являются подкорковыми центрами зрения, оба нижних, coll?culi inferi?res, — подкорковыми центрами слуха. В плоской канавке между верхними бугорками лежит шишковидное тело. Каждый холмик переходит в так называемую ручку холмика, Ьг?chium coll?culi, направляющуюся латерально, кпереди и кверху, к промежуточному мозгу. Ручка верхнего холмика, br?chium coll?culi superi?ris, идет под подушкой, p?lvinar, таламуса к латеральному коленчатому телу, c?rpus genicul?tum later?le. Ручка нижнего холмика br?chium coll?culi inferi?ris, проходя вдоль верхнего края trig?num lemn?sci до s?lcus later?lis mesenc?phali, исчезает под медиальным коленчатым телом, c?rpus genicul?tum medi?le. Названные коленчатые тела относятся уже к промежуточному мозгу.
2. Вентральная часть, ножки мозга, ped?nculi c?rebri, содержит все проводящие пути к переднему мозгу.
Ножки мозга имеют вид двух толстых полуцилиндрических белых тяжей, которые расходятся от края моста под углом и погружаются в толщу полушарий большого мозга.
3. Полость среднего мозга, являющаяся остатком первичной полости среднего мозгового пузыря, имеет вид узкого канала и называется водопроводом мозга, aqued?ctus c?rebri. Он представляет узкий, выстланный эпендимой канал 1,5–2,0 см длиной, соединяющий IV желудочек с III. Дорсально водопровод ограничивается крышей среднего мозга, вентрально — покрышкой ножек мозга.
Внутреннее строение среднего мозга. На поперечном разрезе среднего мозга различают три основные части: I) пластинку крыши, l?mina t?cti; 2) покрышку, tegm?ntum, представляющую верхний отдел ped?nculi c?rebri; 3) вентральный отдел ped?nculi c?rebri, или основание ножки мозга, b?sis ped?nculi cerebr?lis. Соответственно развитию среднего мозга под влиянием зрительного рецептора в нем заложены различные ядра, имеющие отношение к иннервации глаза.
У низших позвоночных верхнее двухолмие служит главным местом окончания зрительного нерва и является главным зрительным центром. У млекопитающих и у человека с переносом зрительных центров в передний мозг остающаяся связь зрительного нерва с верхним холмиком имеет значение только для рефлексов. В ядре нижнего холмика, а также в медиальном коленчатом теле оканчиваются волокна слуховой петли (lemn?scus later?lis). Крыша среднего мозга имеет двустороннюю связь со спинным мозгом — tr?ctus spinotect?lis и tr?ctus tectobulb?ris et tectospin?lis. Последние после перекреста в покрышке идут к мышечным ядрам в продолговатом и спинном мозге. Это так называемый зрительно-звуковой рефлекторный путь, о котором говорилось при описании спинного мозга. Таким образом, пластинку крыши среднего мозга можно рассматривать как рефлекторный центр для различного рода движений, возникающих главным образом под влиянием зрительных и слуховых раздражений.
Водопровод мозга окружен центральным серым веществом, имеющим по своей функции отношение к вегетативной системе. В нем, под вентральной стенкой водопровода, в покрышке ножки мозга заложены ядра двух двигательных черепных нервов — n. oculomot?rius (III пара) на уровне верхнего двухолмия и n. trochle?ris (IV пара) на уровне нижнего двухолмия. Ядро глазодвигательного нерва состоит из нескольких отделов соответственно иннервации нескольких мышц глазного яблока. Медиально и кзади от него помещается еще небольшое, тоже парное, вегетативное добавочное ядро, n?cleus access?rius, и непарное срединное ядро. Добавочное ядро и непарное срединное ядро иннервируют непроизвольные мышцы глаза, m. cili?ris и m. sph?ncter pupillae. Эта часть глазодвигательного нерва относится к парасимпатической системе. Выше (ростральнее) ядра глазодвигательного нерва в покрышке ножки мозга располагается ядро медиального продольного пучка.
Латерально от водопровода мозга находится ядро среднемозгового тракта тройничного нерва, n?cleus mesenceph?licus n. trig?mini.
Ножки мозга делятся, как уже отмечалось, на вентральную часть, или основание ножки мозга, b?sis ped?nculi c?rebralis, и покрышку, tegm?ntum. Границей между ними служит черное вещество, subst?ntia nigra, обязанное своим цветом содержащемуся в составляющих его нервных клетках черному пигменту — меланину (рис. 284).
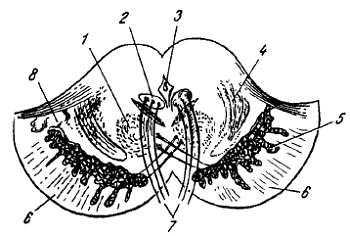
Рис. 284.
1 — nucl. ruber; 2 — nucl. n. oculomotorii; 3 — aqueductus cerebri; 4 — lemniscus medialis; 5 — substantia nigra; 6 — basis pedunculi cerebralis; 7 — n. oculomotorius; 8 — tegmentum mcseneephali.
Покрышка среднего мозга, tegm?ntum mesenc?phali, — часть среднего мозга, расположенная между его крышей и черным веществом (subst?ntia n?gra) ножек мозга.
От нее отходит tr?ctus tegment?lis centr?lis — центральный покрышечный путь — проекционный нисходящий нервный путь, расположенный в центральной части покрышки среднего мозга. Он содержит волокна, идущие от таламуса, бледного шара, красного ядра и ретикулярной формации среднего мозга к ретикулярной формации и оливе продолговатого мозга; относится к экстрапирамидной системе.
Subst?ntia n?gra простирается на всем протяжении ножки мозга от моста до промежуточного мозга; по своей функции относится к экстрапирамидной системе.
Расположенное вентрально от subst?ntia n?gra основание ножки мозга содержит продольные нервные волокна, спускающиеся от коры полушария большого мозга ко всем нижележащим отделам центральной нервной системы (tr?ctus corticopont?nus, corticonucl?aris, corticospin?lis и др.). Tegm?ntum, находящаяся дорсально от subst?ntia n?gra, содержит преимущественно восходящие волокна, в том числе медиальную и латеральную петли. В составе этих петель восходят к большому мозгу все чувствительные пути, за исключением зрительного и обонятельного.
Среди ядер серого вещества самое значительное — красное ядро, n?cleus r?ber. Это удлиненное колбасовидное образование простирается в покрышке ножки мозга от гипоталамуса промежуточного мозга до нижнего двухолмия, где от него начинается важный нисходящий тракт, tr?ctus rubrospin?lis, соединяющий красное ядро с передними рогами спинного мозга. Пучок этот после выхода из красного ядра перекрещивается с аналогичным пучком противоположной стороны в вентральной части срединного шва — вентральный перекрест покрышки. N?cleus r?ber является весьма важным координационным центром экстрапирамидной системы, связанным с остальными ее частями. К нему проходят волокна от мозжечка в составе верхних ножек последнего после их перекреста под крышей среднего мозга, вентрально от aqued?ctus c?rebri, а также от p?llidum — самого нижнего и самого древнего из подкорковых узлов головного мозга, входящих в состав экстрапирамидной системы. Благодаря этим связям мозжечок и экстрапирамидная система через посредство красного ядра и отходящего от него tractus rubrospinalis оказывают влияние на всю скелетную мускулатуру в смысле регуляции бессознательных автоматических движений.
В покрышку среднего мозга продолжаются такжеретикулярная формация, form?tio reticul?ris, и fasciculus longitudin?lis medi?lis. Последний берет начало в различных местах. Одна из его частей начинается из вестибулярных ядер, проходит на той и другой стороне по бокам средней линии, непосредственно под серым веществом дна водопровода и IV желудочка, и состоит из восходящих и нисходящих волокон, идущих к ядрам III, IV, VI и XI черепных нервов. Медиальный продольный пучок является важным ассоциативным путем, связующим различные ядра нервов глазных мышц между собой, чем обусловливаются сочетанные движения глаз при отклонении их в ту или другую сторону. Функция его связана также с движениями глаз и головы, возникающими при раздражении аппарата равновесия.
Передний мозг
Передний мозг, prosenc?phalon, развивается в связи с обонятельным рецептором и вначале (у водных животных) является чисто обонятельным мозгом, rhinenc?phalon. С переходом животных из водной среды в воздушную роль обонятельного рецептора возрастает, так как с его помощью определяются содержащиеся в воздухе химические вещества, сигнализирующие животному о добыче, опасности и других жизненно важных явлениях природы с далекого расстояния, — дистантный рецептор.
Поэтому, а также благодаря развитию и совершенствованию других анализаторов передний мозг у наземных животных сильно разрастается и превосходит другие отделы центральной нервной системы, превращаясь из обонятельного мозга в орган, управляющий всем поведением животного. Соответственно двум основным формам поведения: 1) инстинктивному, основанному на опыте вида (безусловные рефлексы), и 2) индивидуальному, основанному на опыте индивида (условные рефлексы), в переднем мозге развиваются две группы центров: 1) базальные, или подкорковые, ядра полушарий большого мозга; 2) кора большого мозга. В эти две группы центров переднего мозга поступают все нервные импульсы и к ним протягиваются все афферентные чувствительные пути, которые (за немногими исключениями) предварительно проходят через один общий центр — таламус, th?lamus. Приспособление организма к среде путем изменения обмена веществ обусловило возникновение в переднем мозге высших центров, ведающих вегетативными процессами (гипоталамус, hypoth?lamus).
Из двух частей переднего мозга, промежуточного мозга, dicnc?phalon, и конечного, telenc?phalon, кора и подкорковые ядра относятся к конечному мозгу, а таламус и гипоталамус — к промежуточному.
Промежуточный мозг
Промежуточный мозг, dienc?phalon, залегает под мозолистым телом и сводом, срастаясь по бокам с полушариями конечного мозга. Соответственно сказанному выше о функции и развитии переднего мозга в промежуточном мозге различают две основные части: 1) дорсальную (филогенетически более молодую) — thalamenc?phalon — центр афферентных путей и 2) вентральную (филогенетически более старую) — hypoth?lamus — высший вегетативный центр. Полостью dienc?phalon является III желудочек.
Таламический мозг
Thalamenc?phalon в свою очередь состоит из трех частей: th?lamus — таламус, epith?lamus — над таламическая область и metath?lamus — заталамическая область (рис. 285).
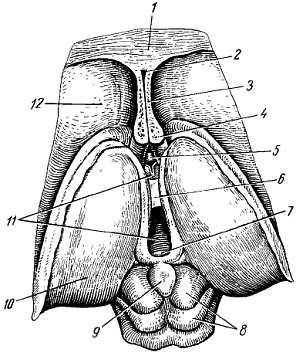
Рис. 285.
1 — corpus callosum; 2 — cavum sepli pellucidi; 3 — septum pellucidum; 4 — fornix (поперечный разрез столбов); 5 — comissura anterior; 6 — adhesio interthalamica; 7 — comissura posterior; 8 — tectum mesencephali (lam. tecti); 9 — corpus pineale; 10 — thalamus; 11 — ventriculus tertius; 12 — nucl. caudatus (caput).
A. Th?lamus, таламус, представляет собой большое парное скопление серого вещества в боковых стенках промежуточного мозга по бокам III желудочка, имеющее яйцевидную форму, причем передний его конец заострен в виде tuberculum ant?rius, а задний расширен и утолщен в виде подушки, р?lvinar. Деление на передний конец и подушку соответствует функциональному делению th?lamus на центры афферентных путей (передний конец) и на зрительный центр (задний). Дорсальная поверхность покрыта тонким слоем белого вещества — str?tum zon?le. В латеральном своем отделе она обращена в полость бокового желудочка, отделяясь от соседнего с ней хвостатого ядра пограничной бороздкой, s?lcus termin?lis, являющейся границей между telenc?phalon, к которому принадлежит хвостатое ядро, и dienc?phalon, к которому относится таламус. По этой бороздке проходит полоска мозгового вещества, str?a termin?lis.
Медиальная поверхность таламуса, покрытая тонким слоем серого вещества, расположена вертикально и обращена в полость III желудочка, образуя его латеральную стенку: Сверху она отграничивается от дорсальной поверхности посредством белой мозговой полоски, str?a medull?ris th?lami. Обе медиальные поверхности таламусов соединены между собой серой спайкой — adh?sio interth?lamica, лежащей почти посередине. Латеральная поверхность таламуса граничит с внутренней капсулой, c?psula int?rna. Нижней своей поверхностью таламус располагается над ножкой мозга, срастаясь с ее покрышкой (рис. 286).
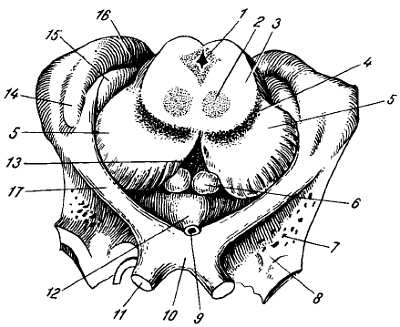
Рис. 286.
1 — aqueductus cerebri; 2 — nucl. ruber; 3 — tegmentum; 4 — substantia nigra; 5 — pedunculus cerebri; 6 — corpus mamillare; 7 — substantia perforata anterior; 8 — trigonum olfactorium; 9 — infundibulum; 10 — chiasma opticum; 11 — n. opticus; 12 — tuber cinereum; 13 — substantia perforata posterior; 14 — corpus geniculatum IateraIe; 15 — corpus gcniculatum mediale; 16 — pulvinar; 17 — tr. opticus.
Как видно на разрезах, серая масса таламуса белыми прослойками, l?minae medull?res th?lami, разделяется на отдельные ядра, носящие названия в зависимости от их топографии: передние, центральные, медиальные, латеральные, вентральные и задние.
Функциональное значение таламуса очень велико. В нем переключаются афферентные пути: в его подушке, p?lvinar, где находится заднее ядро, оканчивается часть волокон зрительного тракта (подкорковый центр зрения, ассоциативное ядро таламуса), в передних ядрах — пучок, идущий от c?rpora mamill?ria и связывающий таламус с обонятельной сферой, и, наконец, все остальные афферентные чувствительные пути от нижележащих отделов центральной нервной системы в остальных его ядрах, причем lemn?scus medi?lis заканчивается в латеральных ядрах. Таким образом, th?lamus является подкорковым центром почти всех видов чувствительности. Отсюда чувствительные пути идут частью в подкорковые ядра (благодаря чему таламус является чувствительным центром экстрапирамидной системы), частью — непосредственно в кору (tr?ctus thalamocortic?lis).
Б. Epith?lamus. Striae medull?res обоих таламусов направляются кзади (каудально) и образуют на той и другой стороне треугольное расширение, называемое trig?num hab?nulae. От последнего отходит так называемый поводок, hab?nula, который вместе с таким же поводком противоположной стороны соединяется с шишковидным телом, c?rpus pine?le (см. рис. 285). Спереди от c?rpus pine?le оба поводка связаны вместе посредством commiss?ra habenul?rum. Само шишковидное тело, напоминающее несколько сосновую шишку (p?nus — сосна, отчего и происходит его название), по своему строению и функции относится к железам внутренней секреции. Выдаваясь кзади в область среднего мозга, шишковидное тело располагается в бороздке между верхними холмиками крыши среднего мозга, образуя как бы пятый бугорок.
В. Metath?lamus. Позади таламуса находятся два небольших возвышения — коленчатые тела, c?rpus genicul?tun later?le et medi?le (см. рис. 278, 286).
Медиальное коленчатое тело, меньшее по размерам, но более выраженное, лежит спереди ручки нижнего холмика под p?lvinar таламуса, отделенное от него ясной бороздкой. В нем заканчиваются волокна слуховой петли, lemn?scus later?lis, вследствие чего оно является вместе с нижними холмиками крыши среднего мозга подкорковым центром слуха. Латеральное коленчатое тело, большее, в виде плоского бугорка помещается на нижней латеральной стороне p?lvinar. В нем оканчивается большей своей частью латеральная часть зрительного тракта (другая часть тракта оканчивается в p?lvinar). Поэтому вместе с p?lvinar и верхними холмиками крыши среднего мозга латеральное коленчатое тело является подкорковым цен гром зрения. Ядра обоих коленчатых тел центральными путями связаны с корковыми концами соответственных анализаторов.
Гипоталамус
Гипоталамус, hypoth?lamus (см. рис. 286), в широком смысле слова, объединяет образования, расположенные вентрально под дном III желудочка, впереди subst?ntia perfor?ta post?rior, включая и заднюю гипоталамическую область, r?gio hypothal?mica post?rior. Соответственно эмбриональному развитию hypothalamus делится на два отдела: передний — r?gio hypothal?mica ant?rior, под именем которого объединяют t?ber ciner?um с infund?bulum и hyp?physis, а также chi?sma ?pticum с tr?ctus ?pticus, задний — c?rpora mamill?ria и regio hypothal?mica post?rior. Ядра гиноталамической области связаны с гипофизом посредством портальных сосудов (с передней долей гипофиза) и гипоталамогипофизарного пучка (с задней долей его).
Благодаря этим связям гипоталамус и гипофиз образуют особую гипоталамо-гипофизарную систему (ГГНС).
A. T?ber ciner?um, серый бугор, находящийся спереди от c?rpora mamill?ria, представляет непарный полый выступ нижней стенки III желудочка, состоящий из тонкой пластинки серого вещества. Верхушка бугра вытянута в узкую полую воронку, infund?bulum, на слепом конце которой находится гипофиз, hyp?physis (gl?ndula pituit?ria), лежащий в углублении турецкого седла (описание его см. в разделе «Органы внутренней секреции»), В t?ber c?nereum заложены ядра серого вещества, являющиеся высшими вегетативными центрами, влияющими, в частности, на обмен веществ и теплорегуляцию.
Б. Chi?sma ?pticurn, зрительный перекрест, лежит впереди серого бугра, образован перекрестом зрительных нервов, nn. ?ptici.
В. C?rpora mamill?ria, сосцевидные тела, — два небольших белого цвета возвышения неправильной шаровидной формы, лежащих симметрично по бокам средней линии, спереди от subst?ntia perfor?ta post?rior. Под поверхностным слоем белого вещества внутри каждого из тел находится два серых ядра.
По своей функции corpora mamill?ria относятся к подкорковым обонятельным центрам.
Г. R?gio hypothal?mica post?rior, задняя гипоталамическая область; это небольшой участок мозгового вещества, расположенный под таламусом. В нем латеральнее subst?ntia n?gra залегает принадлежащее промежуточному мозгу овальное тело, n?cleus hypothal?micus post?rior. Оно является одним из звеньев экстрапирамидной системы; ему также приписывают и вегетативные функции.
III желудочек
III желудочек, ventr?culus t?rtius, расположен как раз по средней линии и на фронтальном разрезе мозга имеет вид узкой вертикальной щели. Боковые стенки III желудочка образованы медиальными поверхностями таламусов, между которыми почти посередине перекидывается adh?sio interthal?mica (см. рис. 285). Переднюю стенку желудочка составляет снизу тонкая пластинка, l?mina termin?lis, а дальше кверху — столбики свода (c?lumnae f?rnicis) с лежащей поперек белой передней спайкой, commiss?re c?rebri ant?rior. По бокам у передней стенки желудочка столбики свода вместе с передними концами таламусов ограничивают межжелудочковые отверстия, for?mina intervetricul?ria, соединяющие полость III желудочка с боковыми желудочками, залегающими в полушариях конечного мозга. Верхняя стенка III желудочка, лежащая под сводом и мозолистым телом, представляет собой t?la choro?dea ventr?culi t?rtii (рис. 287); в состав последней входят недоразвитая стенка мозгового пузыря в виде эпителиальной пластинки, l?mina epitheli?lis, и сросшаяся с ней мягкая оболочка.
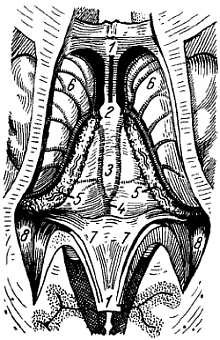
Рис. 287.
1 — corpus callosum; 2 — columnae fornicis (перерезаны); 3 — tela cnoroidea ventriculi tertii; 4 — v. cerebri magna; 5 — plexus choroideus ventriculi lateralis; 6 — nucll. caudati; 7 — crus fornicis; 8 — cornu posterius ventriculi lateralis.
По бокам от средней линии в t?la chorio?dea заложено сосудистое сплетение, pl?xus choroideus ventr?culi t?rtii. В области задней стенки желудочка находятся commiss?ra habenul?rum и commiss?ra c?rebri post?rior, между которыми вдается в каудальную сторону слепой выступ желудочка, rec?ssus pine?lis. Вентрально от commiss?ra post?rior открывается в III желудочек воронкообразным отверстием водопровод. Нижняя, узкая, стенка III желудочка, отграниченная изнутри от боковых стенок бороздками (s?lci hypothal?mici), со стороны основания мозга соответствует subst?ntia perfor?ta post?rior, c?rpora mamill?ria, t?ber ciner?um с chi?sma ?pticum. В области дна полость желудочка образует два углубления: rec?ssus infund?buli, вдающийся в серый бугор и в воронку, и rec?ssus ?pticus, лежащий впереди хиазмы. Внутренняя поверхность стенок III желудочка покрыта эпендимой.
Рассмотренные отделы мозга: ромбовидный (кроме мозжечка), средний и промежуточный — объединяются под названием мозгового ствола. Клиницисты иногда к мозговому стволу относят только задний и средний мозг. Мозговой ствол, филогенетически более старое образование, существенно отличается по строению и функции от более молодой части головного мозга — конечного мозга, telenc?phalon.
Конечный мозг
Как уже отмечалось, конечный мозг, telenc?phalon, представлен двумя полушариями, hemisph?rici c?rebri. В состав каждого полушария входят: плащ, или мантия, p?llium, обонятельный мозг, rhinenc?phalon, и базальные ядра. Остатком первоначальных полостей обоих пузырей конечного мозга являются боковые желудочки, ventr?culi later?les. Передний мозг, из которого выделяется конечный, вначале возникает в связи с обонятельным рецептором (обонятельный мозг), а затем он становится органом управления поведением животного, причем в нем возникают центры инстинктивного поведения, основанного на видовых реакциях (безусловные рефлексы), — подкорковые ядра и центры индивидуального поведения, основанного на индивидуальном опыте (условные рефлексы), — кора большого мозга. Соответственно этому в конечном мозге различают в порядке исторического развития следующие группы центров:
1. Обонятельный мозг, rhinenc?phalon, — самая древняя и вместе с тем самая меньшая часть, расположенная вентрально.
2. Базальные, или центральные, ядра полушарий, «подкорка», — старая часть конечного мозга, paleenc?phalon, скрытая в глубине.
3. Серое вещество коры, c?rtex, — самая молодая часть, neenc?phalon, и вместе с тем самая большая часть, покрывающая остальные как бы плащом, откуда и ее название «плащ», или мантия, p?llium.
Кроме отмеченных для животных двух форм поведения, у человека возникает третья форма — коллективное поведение, основанное на опыте человеческого коллектива, создающегося в процессе трудовой деятельности человека и общения людей с помощью речи. Эта форма поведения связана с развитием самых молодых поверхностных слоев мозговой коры, составляющих материальный субстрат так называемой второй сигнальной (словесной) системы действительности (И. П. Павлов).
Так как в процессе эволюции из всех отделов центральной нервной системы быстрее и сильнее всего растет конечный мозг, то он у человека становится самой большой частью головного мозга и приобретает вид двух объемистых полушарий — правого и левого, hemisph?ria d?xtrum et sin?strum. В глубине продольной щели мозга оба полушария соединены между собой толстой горизонтальной пластинкой — мозолистым телом, c?rpus call?sum, которое состоит из нервных волокон, идущих поперечно из одного полушария в другое. В мозолистом теле различают передний загибающийся книзу конец, иликолено, g?nu c?rporis call?si, среднюю часть, тело, tr?ncus c?rporis call?si, и затем задний конец, утолщенный в форме валика, spl?nium c?rporis call?si. Все эти части хорошо видны на сагиттальном разрезе мозга между обоими полушариями (см. рис. 282). Колено мозолистого тела, загибаясь книзу, заостряется и образует клюв, r?strum c?rporis call?si, который переходит в тонкую пластинку, l?mina rostr?lis, продолжающуюся в свою очередь в l?mina termin?lis.
Под мозолистым телом находится так называемый свод, f?rnix (см. рис. 282; рис. 288), представляющий два дугообразных белых тяжа, которые в средней своей части, c?rpus f?rnicis, соединены между собой, а спереди и сзади расходятся, образуя впереди столбы свода, соl?mпае f?rnicis, позади — ножки свода, cr?ra f?rnicis.
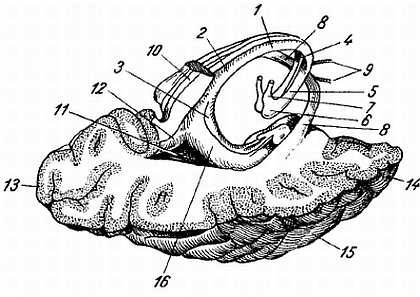
Рис. 288.
1 — corpus fornicis; 2 — crus fornicis; 3 — fimbria hippocampi; 4, 5 — columna fornicis; 6 — corpus mamillare; 7 — fasc. thalamomamillaris; 8, 9 — comissura anterior; 10 — splenium corporis callosi; 11 — trigonum collatcralc; 12 — calcar avis; 13 — polus occipitalis; 14 — polus temporalis; 15 — gyrus temporalis medius; 16 — hippocampus.
Cr?ra f?rnicis, направляясь назад, спускаются в нижние рога боковых желудочков и переходят там в f?mbria hippoc?mpi. Между cr?ra f?rnicis под spl?nium c?rporis call?si протягиваются поперечные пучки нервных волокон, образующие commiss?ra f?rnicis. Передние концы свода, col?nmae f?rnicis, продолжаются вниз до основания мозга, где оканчиваются в c?rpora mamill?ria, проходя через серое вещество hypothalamus. Col?mnae f?rnicis ограничивают лежащие позади них межжелудочковые отверстия, соединяющие III желудочек с боковыми желудочками. Впереди столбов свода находится передняя спайка, commiss?ra ant?rior, имеющая вид белой поперечной перекладины, состоящей из нервных волокон. Между передней частью свода и g?nu c?rporis call?si натянута тонкая вертикальная пластинка мозговой ткани — прозрачная перегородка, s?ptum pell?cidum, в толще которой находится небольшая щелевидная полость, cavum s?pti pell?cidi (см. рис. 282, 293).
Для удобства изучения начнем описание частей конечного мозга в порядке, обратном историческому развитию, т. е. с плаща, который закрывает остальные части.
Плащ
В каждом полушарии можно различить три поверхности: верхнелатеральную, медиальную и нижнюю, и три края: верхний, нижний и медиальный, три конца, или полюса: передний полюс, p?lus front?lis, задний, p?lus occipit?lis, и затем p?lus tempor?lis, соответствующий выступу нижней поверхности и отделенный от нее ямкой, f?ssa later?lis c?rebri.
Поверхность полушария (плащ) образована равномерным слоем серого вещества толщиной 1,3–4,5 мм, содержащего нервные клетки. Слой этот, называемый корой большого мозга, c?rtex c?rebri, представляется как бы сложенным в складки, благодаря чему поверхность плаща имеет в высшей степени сложный рисунок, состоящий из чередующихся между собой в различных направлениях борозд и валиков между ними, называемых извилинами[35], g?ri. Величина и форма борозд подвержены значительным индивидуальным колебаниям, вследствие чего не только мозг различных людей, но даже полушария одной и той же особи по рисунку борозд не вполне похожи.
Глубокими постоянными бороздами пользуются для разделения каждого полушария на большие участки, называемые долями, l?bi последние в свою очередь разделяются на дольки и извилины. Долей каждого полушария пять: лобная (l?bus front?lis), теменная (l?bus pariet?lis), височная (l?bus tempor?lis), затылочная (l?bus occipit?lis) и долька, скрытая на дне латеральной борозды, так называемый островок (?nsula).
Верхнелатеральная поверхность полушария разграничена на доли посредством трех борозд (см. рис. 274; рис. 289): латеральной, центральной и верхнего конца теменно-затылочной борозды, которая, находясь на медиальной стороне полушария, образует зарубку на его верхнем краю.
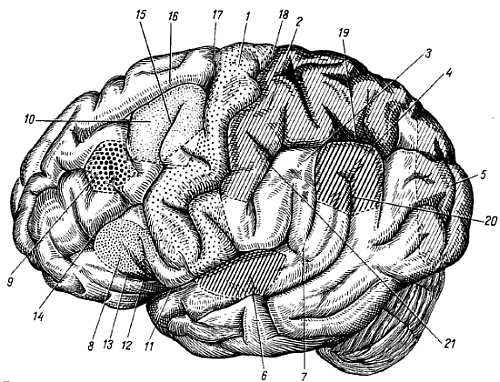
Рис. 289.
1 — ядро двигательного анализатора; 2 — ядро кожного анализатора; 3 — ядро двигательного анализатора, посредством которого синтезируются привычные целенаправленные движения; 4 — ядро зрительного анализатора письменной речи; 5 — ядро зрительного анализатора (зрительная память); 6 — ядро слухового анализатора; 7 — ядро слухового анализатора речи; 8 — ядро двигательного анализатора артикуляции речи; 9 — ядро двигательного анализатора, имеющего отношение к сочетанному повороту головы и глаз; 10 — ядро двигательного анализатора письменной речи; 11, 12, 13 — sul. cerebri lateralis; 11 — сама борозда; 12 — r. ascendens; 13 — r. anterior; 14 — sul. frontalis inferior; 15 — sul. precentralis inferior; 16 — sul. frontalis superior; 17 — sul. precentralis superior; 18 — sul. centralis; 19 — sul. intraparietalis; 20 — sul. temporalis superior; 21 — sul. postcentralis.
Латеральная борозда, s?lcus c?rebri later?lis, начинается на базальной поверхности полушария из латеральной ямки и затем переходит на верхнелатеральную поверхность, направляясь назад и несколько вверх. Она оканчивается приблизительно на границе средней и задней третей верхнелатеральной поверхности полушария. В передней части латеральной борозды от нее отходят две небольшие ветви: r?mus asc?nclens и r?mus ant?rior, направляющиеся в лобную долю.
Центральная борозда, s?lcus centr?lis, начинается на верхнем краю полушария, несколько кзади от его середины, и идет вперед и вниз. Нижний конец центральной борозды не доходит до латеральной борозды. Участок полушария, находящийся впереди центральной борозды, относится к лобной доле; часть мозговой поверхности, лежащая сзади от центральной борозды, составляет теменную долю, которая посредством задней части латеральной борозды отграничивается от лежащей ниже височной доли. Задней границей теменной доли служит конец вышеупомянутой теменно-затылочной борозды, s?lcus parietooccipit?lis, расположенной на медиальной поверхности полушария, но эта граница неполная, ибо названная борозда не заходит далеко на верхнелатеральную поверхность, вследствие чего теменная доля непосредственно переходит в затылочную. Эта последняя также не имеет резкой границы, которая отделяла бы ее от впереди лежащей височной доли. Вследствие этого граница между только что упомянутыми долями проводится искусственно посредством линии, идущей от теменно-затылочной борозды к нижнему краю полушария.
Каждая доля состоит из ряда извилин, называемых в отдельных местах дольками, которые ограничиваются бороздами мозговой поверхности.
Лобная доля. В заднем отделе наружной поверхности этой доли проходит s?lcus precentr?lis почти параллельно направлению s?lcus centr?lis. От нее в продольном направлении отходят две борозды: s?lcus front?lis sup?rior et s?lcus front?lis inf?rior. Благодаря этому лобная доля разделяется на четыре извилины — одну вертикальную и три горизонтальные. Вертикальная извилина, g?rus precentr?lis, находится между s?lcus centr?lis и s?lcus precentr?lis.
Горизонтальные извилины лобной доли следующие: 1) верхняя лобная, g?rus front?lis sup?rior, которая идет выше s?lcus front?lis sup?rior, параллельно верхнему краю полушария, заходя и на его медиальную поверхность; 2) средняя лобная извилина, g?rus front?lis m?dius, тянется между верхней и нижней лобными бороздами и 3) нижняя лобная извилина, g?rus front?lis inf?rior, помещается между s?lcus front?lis inf?rior и латеральной бороздой. Ветви латеральной борозды, вдающиеся в нижнюю лобную извилину, делят последнюю на три части: pars opercul?ris, лежащую между нижним концом s?lcus precentr?lis и r?mus asc?ndens s?lci later?lis, pars triangul?ris, находящуюся между обеими ветвями латеральной борозды, и, наконец, pars orbit?lis, помещающуюся впереди от r?mus ant?rior s?lci later?lis.
Теменная доля. На ней приблизительно параллельно центральной борозде располагается s?lcus postcentr?lis, сливающаяся обычно с s?lcus intrapariet?lis, идущей в горизонтальном направлении. В зависимости от расположения этих борозд теменная доля разделяется на три извилины, из которых одна вертикальная, а две другие горизонтальные. Вертикальная извилина, g?rus postcentr?lis, идет позади s?lcus centr?lis в одном направлении с g?rus precentr?lis, отделенная от нее центральной бороздой. Выше s?lcus intrapariet?lis помещается верхняя теменная извилина, или долька, l?bulus pariet?lis sup?rior, которая распространяется и на медиальную поверхность полушария. Ниже s?lcus intrapariet?lis лежит l?bulus pariet?lis inf?rior, которая, направляясь назад, огибает концы латеральной борозды и s?lcus tempor?lis sup?rior и теряется в области затылочной доли. Часть l?bulus pariet?lis inf?rior, огибающая латеральную борозду, называется g?rus supramargin?lis; другая часть, которая огибает s?lcus tempor?lis sup?rior, носит название g?rus angul?ris.
Височная доля. Латеральная поверхность этой доли имеет три продольные извилины, отграниченные друг от друга s?lcus tempor?lis sup?rior и s?lcus tempor?lis inf?rior. Верхняя из извилин, g?rus tempor?lis sup?rior, находится между латеральной бороздой и s?lcus tempor?lis sup?rior. Верхняя ее поверхность, скрытая в глубине латеральной борозды, несет 2–3 короткие извилинки, называемые g?ri temp?rales transversi. Между верхней и нижней височными бороздами протягивается gyrus tempor?lis m?dius. Ниже последней, отделяясь от нее s?lcus tempor?lis inf?rior, проходит g?rus tempor?lis inf?rior, которая посредством нижнего края отделена от лежащей на нижней поверхности g?rus occipitotempor?lis later?lis.
Затылочная доля. Борозды латеральной поверхности этой доли изменчивы и непостоянны. Из них выделяют идущую поперечно s?lcus occipit?lis transv?rsus, соединяющуюся обыкновенно с концом s?lcus intrapariet?lis.
Островок, ?nsula. Чтобы увидеть эту дольку, надо раздвинуть или удалить нависающие над ней края латеральной борозды. Эти края, относясь к лобной, теменной и височной долям, носят название покрышки, op?rculum. Островок имеет форму треугольника, верхушка которого обращена вперед и вниз. Спереди, сверху и сзади островок отграничивается от соседних с ним частей посредством глубокой борозды, s?lcus circul?ris. Поверхность островка покрыта короткими извилинами.
Нижняя поверхность полушария (см. рис. 276) в той ее части, которая лежит кпереди от латеральной ямки, относится к лобной доле. Здесь параллельно медиальному краю полушария проходит s?lcus olfact?rius, в которой лежат b?lbus et tr?ctus olfact?rius. Между этой бороздой и медиальным краем полушария протягивается прямая извилина, g?rus r?ctus, представляющая собой продолжение верхней лобной извилины. Латерально от s?lcus olfact?rius на нижней поверхности находится несколько непостоянных бороздок, s?lci orbit?les, ограничивающих g?ri orbit?les, которые можно рассматривать как продолжение средней и нижней лобных извилин. Задний участок базальной поверхности полушария образован нижними поверхностями височной и затылочной долей, которые здесь не имеют определенных границ. На этом участке видны две борозды: s?lcus occipitotempor?lis, проходящая в направлении от затылочного полюса к височному и ограничивающая g?rus occipitotempor?lis later?lis, и идущая параллельно ей s?lcus collater?lis (продолжением ее кпереди является s?lcus rhin?lis). Между ними располагается g?rus occipitotempor?lis medi?lis. Медиально от s?lcus collater?lis расположены две извилины: между задним отделом этой борозды и sulcus calcarinus лежит g?rus lingu?lis; между передним отделом этой борозды и s?lcus rhin?lis, с одной стороны, и глубокой s?lcus hippoc?mpi, огибающей ствол мозга, — с другой лежит g?rus parahippocamp?lis. Эта извилина, примыкающая к стволу мозга, находится уже на медиальной поверхности полушария.
Медиальная поверхность полушария. На этой поверхности (см. рис. 282) находится борозда мозолистого тела, s?lcus c?rporis call?si, идущая непосредственно над мозолистым телом и продолжающаяся своим задним концом в глубокую s?lcus hippoc?mpi, которая направляется вперед и книзу. Параллельно и выше этой борозды проходит по медиальной поверхности полушария s?lcus c?nguli, которая начинается спереди под клювом мозолистого тела, затем идет назад и оканчивается своим задним концом на верхнем краю полушария. Пространство, располагающееся между этим краем полушария и s?lcus c?nguli, относится к лобной доле, к верхней лобной извилине. Небольшой участок над s?lcus c?nguli, ограниченный сзади задним концом s?lcus c?nguli, а спереди маленькой бороздкой, s?lcus paracentr?lis, называется парацентральной долькой, l?bulus paracentr?lis, так как он соответствует медиальной поверхности верхних концов обеих центральных извилин, переходящих здесь друг в друга.
Кзади от l?bulus paracentr?lis находится четырехугольная поверхность (так называемое предклинье, prec?neus), ограниченная спереди концом s?lcus c?nguli, снизу небольшой s?lcus subpariet?lis, а сзади глубокой s?lcus parietoocipit?lis. Prec?neus относится к теменной доле. Позади prec?neus лежит резко обособленный участок коры, относящийся к затылочной доле, — клин, c?neus, который ограничен спереди s?lcus parietooccipit?lis, а сзади s?lcus calcar?nus, сходящимися под углом. Книзу и кзади клин соприкасается с g?rus lingu?lis. Между s?lcus c?nguli и бороздой мозолистого тела протягивается поясная извилина, g?rus c?nguli, которая при посредстве перешейка, ?sthmus, продолжается в g?rus parahippocamp?lis, заканчивающуюся крючком, ?ncus. Парагиппокампальная извилина ограничивается с одной стороны s?lcus hippoc?mpi, огибающей ствол мозга, а с другой — s?lcus cojlater?lis и ее продолжением кпереди, носящим название s?lcus rhin?lis. ?sthmus — суженное место перехода поясной извилины в парагиппокампальную, находится позади spl?nium c?rporis call?si, у конца борозды, образовавшейся от слияния s?lcus parietooccipit?lis с s?lcus calcar?nus. G?rus c?nguli, ?sthmus и g?rus parahippocamp?lis образуют вместе сводчатую извилину, g?rus fornic?tus, которая описывает почти полный круг, открытый только снизу и спереди. Сводчатая извилина не имеет отношения ни к одной из долей плаща. Она относится к лимбической области.
Лимбическая область (r?gie l?mbica) — часть новой коры полушарий большого мозга, занимающая поясную и нарагиппокампальную извилины; входит в состав лимбической системы. Раздвигая край s?lcus hippoc?mpi, можно видеть узкую зазубренную серую полоску, представляющую собой рудиментарную извилину g?rus dentatus.
Строение мозговой коры. Кора полушарий большого мозга состоит из шести слоев (пластинок), различающихся между собой главным образом по форме входящих в них нервных клеток (рис. 290): 1) молекулярная пластинка лежит непосредственно под pia mater и содержит концевые разветвления отростков нервных клеток, переплетающихся сетеобразно; 2) наружная зернистая пластинка называется так потому, что в ее состав входят многочисленные маленькие клетки, похожие на зерна; 3) наружная пирамидная пластинка состоит из малых и средних пирамидных нервных клеток; 4) внутренняя зернистая пластинка слагается, так же как и наружная зернистая, из маленьких клеток — зерен; 5) внутренняя пирамидная пластинка содержит большие пирамидные клетки; 6) мультиформная пластинка граничит с белым веществом. Из этих 6 слоев нижние (V и VI) являются преимущественно началом эфферентных путей; в частности, V слой состоит из пирамидных клеток, аксоны которых составляют пирамидную систему (пирамидные клетки, дающие начало пирамидной системе, находятся в предцентральной извилине). Средние слои (III и IV) связаны преимущественно с афферентными путями, а верхние (I и II) относятся к ассоциативным путям коры. Шестислойный тип коры видоизменяется в различных областях как в смысле толщины и расположения слоев, так и состава клеток (подробно см. в курсе гистологии).
Рис. 290.
1 — молекулярная пластинка; 2 — наружная зернистая пластинка; 3 — наружная пирамидная пластинка; 4 — внутренняя зернистая пластинка; 5 — внутренняя пирамидная пластинка; 6 — мультиформная пластинка; 7 — белое вещество.
Обонятельный мозг
Обонятельный мозг, rhinenc?phalon (рис. 291, 292), есть филогенетически самая древняя часть переднего мозга, возникшая в связи с анализатором обоняния, когда передний мозг не стал еще органом поведения животного. Поэтому все компоненты его являются различными частями обонятельного анализатора (понятие об анализаторе см. «Морфологические основы локализации функций»).
У рыб почти весь передний мозг является органом обоняния. С развитием новой коры, что наблюдается у млекопитающих и человека, развивается новая часть переднего мозга (neenc?phalon) — плащ, p?llium. Но и плащ проходит свой длинный путь развития и содержит три части различной филогенетической давности. Более старые части.
Рис. 291
а — змеи; б — сумчатого млекопитающего; 1, 3 — neopallium: 2 — archipallium; 4 — hippocampus.
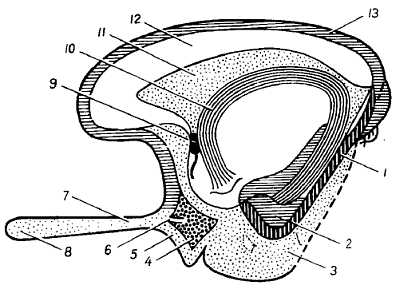
Рис. 292
1 — gyrus dentatus; 2 — gyrus parahippocampalis; 3 — uncus; 4 — substantia perforata anterior; 5, 6 — striae olfactoriae; 7 — tr. olfactorius; 8 — bulbus olfactorius; 9 — commissura anterior; 10 — fornix; 11 — septum pellucidum; 12 — corpus callosum; 13 — gyrus for meatus.
1. Paleop?llium, входящий в состав височной доли. Вначале этот отдел располагался на латеральной поверхности полушария, но в дальнейшем, под влиянием сильно увеличивающегося neop?llium, он свернулся в колбасовидное образование — гиппокамп и сместился медиально в полость бокового желудочка конечного мозга в виде выпячивания его нижнего рога. Гиппокамп покрыт древней корой, paleoc?rtex.
2. Archip?llium — небольшой участок коры на вентральной поверхности лобной доли, лежащий вблизи b?lbus olfact?rius и покрытый старой корой, archic?rtex.
3. Neop?llium, новый плащ, в коре которого, neoc?rtex, появились высшие центры обоняния — корковые концы анализатора. Это — ?ncus, являющийся частью сводчатой извилины.
В результате обонятельный мозг человека содержит ряд образований различного происхождения, которые топографически можно разделить на два отдела. Периферический отдел — это обонятельная доля, l?bus olfact?rius, под которой разумеется ряд образований, лежащих на основании мозга: 1) b?lbus olfact?rius; 2) tr?ctus olfact?rius; 3) trig?num olfact?rium; 4) subst?ntia perfor?ta ant?rior. Центральный отдел — это извилины мозга: 1) парагиппокампальная извилина, g?rus parahippocamp?lis; 2) зубчатая извилина, g?rus dent?tus; 3) сводчатая извилина, g?rus fornic?tus, с расположенной вблизи височного полюса передней ее частью — крючком, ?ncus.
Боковые желудочки
В полушариях конечного мозга залегают ниже уровня мозолистого тела симметрично по сторонам средней линии два боковых желудочка, ventr?culi later?les (рис. 293, 294, 295[рисунка нет]), отделенные от верхнелатеральной поверхности полушарий всей толщей мозгового вещества.
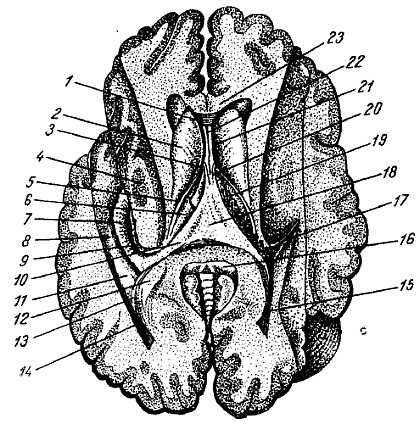
Рис. 293.
1 — cornu anterius; 2 — nucl. caudatus (caput); 3 — for. interventriculare; 4 — nucl. lentiformis (в разрезе); 5 — stria terminalis; 6 — верхняя поверхность thalamus; 7 — hippocampus; 8 — eminentia collateralis; 9 — fimbria hippocampi; 10 — crus fornicis; 11 — cornu posterius ventriculi lateralis; 12 — медиальная стенка заднего рога; 13 — calcar avis; 14, 15 — cornu posterius; 16 — splenium corporis callosi; 17, 19 — plexus choroideus в центральной части бокового желудочка и продолжение его в нижний рог; 18 — commissure fornicis; 20 — columnae fornicis; 21 — septum pellucidum; 22 — cavum septi pellucidi; 23 — corpus callosum.
Полость каждого бокового желудочка (см. рис. 294) соответствует форме полушария: она начинается в лобной доле в виде загнутого вниз и в латеральную сторону переднего рога, c?rnu ant?rius, отсюда она через область теменной доли тянется под названием центральной части, p?rs centr?lis, которая на уровне заднего края мозолистого тела разделяется на нижний рог, c?rni inf?rius, (в толще височной доли) и задний рог, c?rnu post?rius (в затылочной доле).
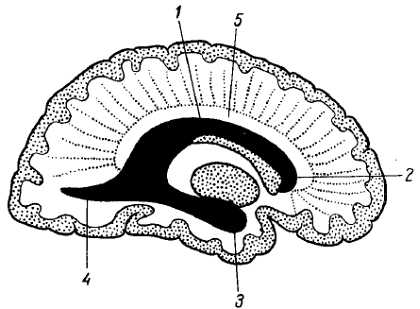
Рис. 294.
1 — pars centralis; 2 — cornu anterius; 3 — cornu inferius; 4 — cornu posterius; 5 — corpus callosum.
Медиальная стенка переднего рога образована s?ptum pell?cidum, которая отделяет передний рог от такого же рога другого полушария (см. рис. 293). Латеральная стенка и отчасти дно переднего рога заняты возвышением серого цвета, головкой хвостатого ядра, c?put n?clei caud?ti, а верхняя стенка образуется волокнами мозолистого тела. Крыша центральной, наиболее узкой части бокового желудочка также состоит из волокон мозолистого тела, дно же составляется из продолжения хвостатого ядра, c?rpus n?clei caud?ti, и части верхней поверхности таламуса. Задний рог окружен слоем белых нервных волокон, происходящих из мозолистого тела, так называемого tap?tum (покров); на его медиальной стенке заметен валик — птичья шпора, c?lcar ?vis, образованная вдавлением со стороны s?lcus calcar?nus, находящейся на медиальной поверхности полушария. Верхнелатеральная стенка нижнего рога образуется tap?tum, составляющим продолжение такого же образования, окружающего задний рог. С медиальной стороны на верхней стенке проходит загибающаяся книзу и кпереди утонченная часть хвостатого ядра — c?uda n?clei caud?ti.
По медиальной стенке нижнего рога на всем протяжении тянется белого цвета возвышение — гиппокамп, hippoc?mpus, который образуется вследствие вдавления от глубоко врезывающейся снаружи s?lcus hippoc?mpi. Передний конец hippoc?mpus разделяется бороздками на несколько небольших бугорков. По медиальному краю гиппокампа идет так называемая бахромка, f?mbria hippoc?mpi, представляющая продолжение ножки свода (crus f?rnicis). На дне нижнего рога находится валик, emin?ntia collater?lis, происходящий от вдавления снаружи одноименной борозды. С медиальной стороны бокового желудочка в его центральную часть и нижний рог вдается мягкая мозговая оболочка, образующая в этом месте сосудистое сплетение, pl?xus choro?deus ventr?culi later?lis. Сплетение покрыто эпителием, представляющим остаток неразвитой медиальной стенки желудочка. Pl?xus choro?deus ventr?culi later?lis является латеральным краем t?la choro?dea ventr?culi t?rtii.
Базальные ядра полушарий
Кроме серой коры на поверхности полушария, имеются еще скопления серого вещества в его толще, именуемые базальными ядрами и составляющие то, что для краткости называют подкоркой. В отличие от коры, имеющей строение экранных центров, подкорковые ядра имеют строение ядерных центров. Различают три скопления подкорковых ядер: c?rpus stri?tum, cl?ustrum и c?rpus amygdalo?deum (рис. 296, 297).
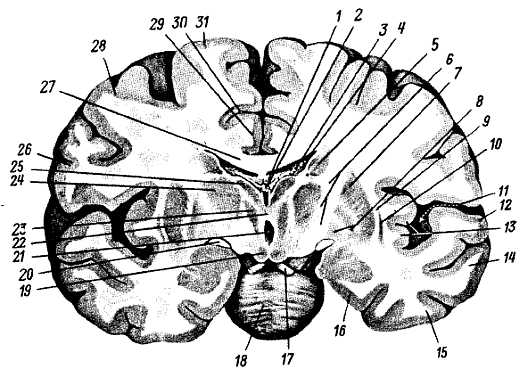
Рис. 296.
1 — fornix; 2 — plexus choroideus ventriculi tertii; 3 — plexus choroideus ventriculi lateralis; 4 — ventriculus lateralis; 5 — nucl. caudatus; 6 — crus posterius внутренней капсулы; 7 — putamen; 8 — globus pallidus; 9 — capsula externa; 10 — claustrum; 11 — sul. cerebri lateralis; 12, 14, 15 — gyri temporales superior, medius et inferior; 13 — insula; 16 — gyrus narahippocampalis; 17 — n. oculomotorius; 18 — pons; 19 — nucl. corporis mamillaris; 20 — tr. opticus; 21 — ventriculus tertius; 22 — adhesio interthalamica; 23, 24, 25 — ядра thalamus; 26, 28, 31 — gyri frontales inferior, medius et superior; 27 — corpus callosum; 29 — gyrus cinguli; 30 — sul. cinguli.
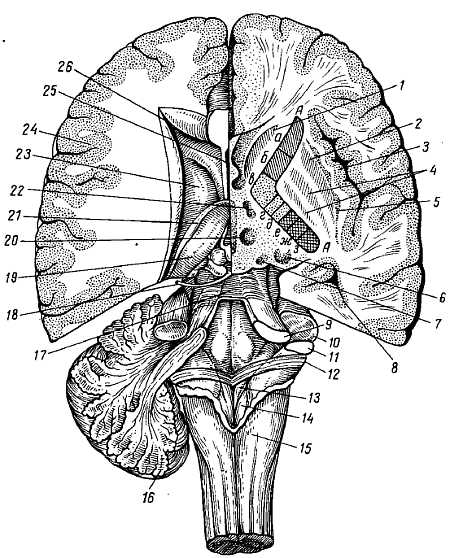
Рис. 297.
1 — nucl. caudatus; 2 — putamen; 3 — кора островка полушария; 4 — globus pallidus; 5 — claustrum; 6 — cauda nuclei caudati; 7 — nucl. corporis geniculati medialis; 8 — cornu inferius ventriculi lateralis; 9 — pedunculus cerebellaris superior; 10 — pedunculus cerebellaris medius; 11 — pedunculus cerebellaris inferior; 12 — striae medullares; 13 — trigonum n. hypoglossi; 14 — trigonum n. vagi; 15 — tuberculum gracile; 16 — cerebellum; 17 — velum medullare superius; 18 — n. trochlearis; 19 — thalamus; 20 — nucl. ruber; 21 — stria terminalis; 22 — nucl. hypothalamicus; 23 — nucl. caudatus; 24 — cortex cerebri; 25 — cavum septi pellucidi; 26 — cornu anterius ventriculi lateralis; AA — capsula interna: a — fibrae corticothalanncae; б — tr. frontopontinus; в — tr. corticonuclearis; г — tr. corticospinalis; д — fibrae thalamocorticales; e — tr. occipitotemporopontinus; ж — центральный слуховой тракт, з — центральный зрительный тракт.
1. C?prus stri?tum, полосатое тело, состоит из двух не вполне отделенных друг от друга частей — n?cleus caud?tus и n?cleus lentif?rmis.
A. N?cleus caud?tus, хвостатое ядро, лежит выше и медиальнее n?cleus lentif?rmis, отделяясь от последнего прослойкой белого вещества, называемой внутренней капсулой, c?psula int?rna. Утолщенная передняя часть хвостатого ядра, его головка, c?put n?clei caud?ti, образует латеральную стенку переднего рога бокового желудочка, задний же утонченный отдел хвостатого ядра, c?rpus et c?uda n?clei caud?ti, тянется назад по дну центральной части бокового желудочка; c?uda заворачивается на верхнюю стенку нижнего рога. С медиальной стороны n?cleus caud?tus прилегает к таламусу, отделяясь от него полоской белого вещества, str?a termin?lis. Спереди и снизу головка хвостатого ядра доходит до subst?ntia perfor?ta ant?rior, где она соединяется с n?cleus lentif?rmis (с частью последнего, называемой put?men). Кроме этого широкого соединения обоих ядер с вентральной стороны, имеются еще тонкие полоски серого вещества, располагающиеся вперемешку с белыми пучками внутренней капсулы. Они послужили причиной названия «полосатое тело», c?rpus stri?tum (см. рис. 296).
Б. N?cleus lentif?rmis, чечевицеобразное ядро, залегает латерально от n?cleus caud?tus и таламуса, отделенное от них c?psula int?rna. На горизонтальном разрезе полушария медиальная поверхность чечевицеобразного ядра, обращенная к внутренней капсуле, имеет форму угла с верхушкой, направленной к середине; передняя сторона угла параллельна хвостатому ядру, а задняя — таламусу. Латеральная поверхность немного выпукла и обращена к латеральной стороне полушария в области островка. Спереди и вентрально, как было уже указано, чечевицеобразное ядро сливается с головкой n?cleus caud?tus. На фронтальном разрезе чечевицеобразное ядро имеет форму клина, верхушка которого обращена в медиальную сторону, а основание — в латеральную. Чечевицеобразное ядро двумя параллельными белыми прослойками, l?minae medull?res, разделяется на три членика, из которых латеральный, темно-серого цвета, называется скорлупой, put?men, а два медиальных, более светлых, носят вместе название бледного шара, gl?bus p?llidus (см. рис. 296).
Отличаясь уже по своему макроскопическому виду, gl?bus p?llidus имеет также и гистологическую структуру, отличную от других частей полосатого тела. Филогенетически gl?bus p?llidus представляет более старое образование (paleostri?tum), чем put?men и n?cleus caud?tus (neostri?tum).
Ввиду всех этих особенностей gl?bus p?llidus в настоящее время выделяют в особую морфологическую единицу под названием p?llidum, тогда как обозначение stri?tum оставляют только за put?men и n?cleus caud?tus. Вследствие этого термин «чечевицеобразное ядро» теряет свое прежнее значение и может употребляться только в чисто топографическом смысле, а вместо прежнего названия c?rpus stri?tum хвостатое и чечевицеобразное ядро именуют стриопаллидарной системой. Стриопаллидарная система представляет собой главную часть экстрапирамидной системы (см. далее), а кроме того, она является высшим регулирующим центром вегетативных функций в отношении теплорегуляции и углеводного обмена, доминирующим над подобными же вегетативными центрами в hypoth?lamus.
2. Cl?ustrum, ограда, представляет тонкую пластинку серого вещества, заложенную в области островка, между ним и put?men (см. рис. 297). От последнего она отделяется прослойкой белого вещества, c?psula ext?rna, а от коры островка — прослойкой, носящей название c?psula extr?ma.
3. C?rpus amygdalo?deum, миндалевидное тело, расположено под put?men в переднем конце височной доли. C?rpus amygdalo?deum, по-видимому, относится к подкорковым обонятельным центрам и к лимбической системе. В нем оканчивается идущий из обонятельной доли и subst?ntia perfor?ta ant?rior пучок волокон, отмеченный при описании таламуса под названием str?a termin?lis (см. рис. 297).
Лимбическая система представляет комплекс образований конечного, промежуточного и среднего мозга, участвующий в регуляции различных вегетативных функций, поддержании постоянства внутренней среды организма (гомеостаза) и в формировании эмоционально окрашенных поведенческих реакций. Поэтому некоторые авторы обозначают лимбическую систему как «висцеральный мозг». Основную часть ее составляют структуры коры большого мозга, расположенные преимущественно на медиальной поверхности его полушарий, и тесно связанные с ними подкорковые образования, а именно: амигдалоидная область, конечная полоска, гипоталамус, гиппокамп, свод, септальная область, сосцевидные тела, сосцевидно-таламический пучок, таламус, поясная извилина. На медиальной поверхности полушарий большого мозга лимбическая система представлена поясной и парагиппокампальной извилинами.
Белое вещество полушарий
Все пространство между серым веществом мозговой коры и базальными ядрами занято белым веществом. Оно состоит из большого количества нервных волокон, идущих в различных направлениях и образующих проводящие пути конечного мозга. Нервные волокна могут быть разделены на три системы: 1) ассоциативные, 2) комиссуральные и 3) проекционные волокна.
А. Ассоциативные волокна (рис. 298) связывают между собой различные участки коры одного и того же полушария. Они разделяются на короткие и длинные. Короткие волокна, f?brae arcu?tae c?rebri, связывают между собой соседние извилины в форме дугообразных пучков. Длинные ассоциативные волокна соединяют более отдаленные друг от друга участки коры. Таких пучков волокон существует несколько. C?ngulum, пояс, — пучок волокон, проходящий в g?rus fornic?tus, соединяет различные участки коры g?rus cinguli как между собой, так и с соседними извилинами медиальной поверхности полушария. Лобная доля соединяется с нижней теменной долькой, затылочной долей и задней частью височной доли посредством fasc?culus longitudin?lis sup?rior. Височная и затылочная доли связываются между собой через fasc?culus longitudin?lis inf?rior. Наконец, орбитальную поверхность лобной доли соединяет с височным полюсом так называемый крючковидный пучок, fasc?culus uncin?tus.
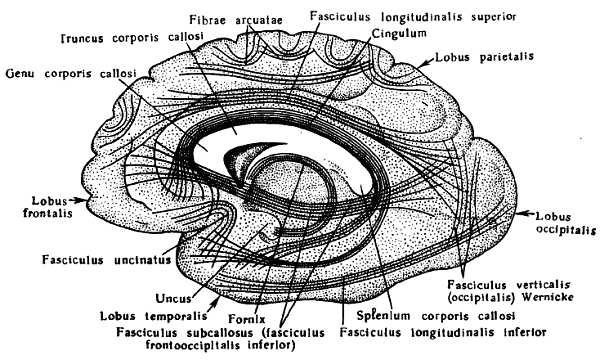
Рис. 298.
Б. Комиссуральные волокна, входящие в состав так называемых мозговых комиссур, или спаек, соединяют симметричные части обоих полушарий. Самая большая мозговая спайка — мозолистое тело, c?rpus call?sum, связывает между собой части обоих полушарий, относящиеся к neenc?phalon.
Две мозговые спайки, commiss?ra ant?rior и commiss?ra f?rnicis, гораздо меньшие по своим размерам, относятся к rhinenc?phalon и соединяют: commiss?ra ant?rior — обонятельные доли и обе парагиппокампальные извилины, commiss?ra forn?cis — гиппокампы.
В. Проекционные волокна связывают мозговую кору частью с th?lamus и c?rpora genicul?ta, частью с нижележащими отделами центральной нервной системы до спинного мозга включительно. Одни из этих волокон проводят возбуждения центростремительно, по направлению к коре, а другие, наоборот, — центробежно.
Проекционные волокна в белом веществе полушария ближе к коре образуют так называемый лучистый венец, cor?na radi?ta, и затем главная часть их сходится во внутреннюю капсулу, о которой упоминалось выше. Внутренняя капсула, c?psula int?rna, как было указано, представляет слой белого вещества между n?cleus lentif?rmis, с одной стороны, и хвостатым ядром и таламусом — с другой. На фронтальном разрезе мозга внутренняя капсула имеет вид косо идущей белой полосы, продолжающейся в ножку мозга. На горизонтальном разрезе она представляется в форме угла, открытого в латеральную сторону (см. рис. 297); вследствие этого в c?psula int?rna различают переднюю ножку, crus ant?rius c?psulae int?rnae, — между хвостатым ядром и передней половиной внутренней поверхности n?cleus lentif?rmis, заднюю ножку, crus post?rius, — между таламусом и задней половиной чечевицеобразного ядра и колено, g?nu c?psulae int?rnae, лежащее на месте перегиба между обеими частями внутренней капсулы. Проекционные волокна по их длине могут быть разделены на следующие системы, начиная с самых длинных:
1. Tr?ctus corticospin?lis (pyramid?lis) проводит двигательные волевые импульсы к мышцам туловища и конечностей. Начавшись от пирамидных клеток коры средней и верхней частей предцентральной извилины и l?bulus paracentr?lis, волокна пирамидного пути идут в составе лучистого венца, а затем проходят через внутреннюю капсулу, занимая передние две трети ее задней ножки, причем волокна для верхней конечности идут спереди волокон для нижней конечности. Далее они проходят через ножку мозга, ped?nculus c?rebri, а оттуда через мост в продолговатый мозг.
2. Tr?ctus corticonucle?ris — проводящие пути к двигательным ядрам черепных нервов. Начавшись от пирамидных клеток коры нижней части предцентральной извилины, они проходят через колено внутренней капсулы и через ножку мозга, затем вступают в мост и, переходя на другую сторону, оканчиваются в двигательных ядрах противоположной стороны, образуя перекрест. Небольшая часть волокон оканчивается без перекреста.
Так как все двигательные волокна собраны на небольшом пространстве во внутренней капсуле (колено и передние две трети задней ножки ее), то при повреждении их в этом месте наблюдается односторонний паралич (hemiplegia) противоположной стороны тела.
3. Tr?tctus corticopont?ni — пути от мозговой коры к ядрам моста. Они идут от коры лобной доли (tr?ctus frontopont?nus), затылочной (tr?ctus occipitopont?nus), височной (tr?ctus temporopont?nus) и теменной (tr?ctus раrietopont?nus). В качестве продолжения этих путей из ядер моста идут волокна в мозжечок в составе его средних ножек. При помощи этих путей кора большого мозга оказывает тормозящее и регулирующее влияние на деятельность мозжечка.
4. F?brae thalamocortic?lis et corticothal?mici — волокна от таламуса к коре и обратно от коры к таламусу. Из волокон, идущих от таламуса, необходимо отметить так называемую центральную таламическую лучистость, которая является конечной частью чувствительного пути, направляющегося к центру кожного чувства в постцентральную извилину. Выходя из латеральных ядер таламуса, волокна этого пути проходят через заднюю ножку внутренней капсулы, позади пирамидного пути. Место это было названо чувствительным перекрестом, так как здесь проходят и другие чувствительные пути, а именно: зрительная лучистость, radi?tio ?ptica, идущая от c?rpus genicul?tum later?le и p?lvinar таламуса к зрительному центру в коре затылочной доли, затем слуховая лучистость, radi?tio ac?stica, направляющаяся от c?rpus genicul?tum medi?le и нижнего холмика крыши среднего мозга к верхней височной извилине, где заложен центр слуха. Зрительный и слуховой пути занимают самое заднее положение в задней ножке внутренней капсулы.
Морфологические основы динамической локализации функций в коре полушарий большого мозга (центры мозговой коры)
Знание локализации функций в коре головного мозга имеет огромное теоретическое значение, так как дает представление о нервной регуляции всех процессов организма и приспособлении его к окружающей среде. Оно имеет и большое практическое значение для диагностики мест поражения в полушариях головного мозга.
Представление о локализации функций в коре головного мозга связано прежде всего с понятием о корковом центре. Еще в 1874 г. киевский анатом В. А. Бец выступил с утверждением, что каждый участок коры отличается по строению от других участков мозга. Этим было положено начало учению о разнокачественности коры головного мозга — цитоархитектонике (цитос — клетка, архитектонес — строю). В настоящее время удалось выявить более 50 различных участков коры — корковых цитоархитектонических полей, каждое из которых отличается от других по строению и расположению нервных элементов. Из этих полей, обозначаемых номерами, составлена специальная карта мозговой коры человека (рис. 299).
Рис. 299.
По И. П. Павлову, центр — это мозговой конец так называемого анализатора. Анализатор — это нервный механизм, функция которого состоит в том, чтобы разлагать известную сложность внешнего и внутреннего мира на отдельные элементы, т. е. производить анализ. Вместе с тем благодаря широким связям с другими анализаторами здесь происходит и синтез, сочетание анализаторов друг с другом и с разными деятельностями организма. «Анализатор есть сложный нервный механизм, начинающийся наружным воспринимающим аппаратом и кончающийся в мозгу» (И. П. Павлов).
С точки зрения И. П. Павлова, мозговой центр, или корковый конец анализатора, имеет не строго очерченные границы, а состоит из ядерной и рассеянной частей — теория ядра и рассеянных элементов. «Ядро» представляет подробную и точную проекцию в коре всех элементов периферического рецептора и является необходимым для осуществления высшего анализа и синтеза. «Рассеянные элементы» находятся по периферии ядра и могут быть разбросаны далеко от него; в них осуществляются более простой и элементарный анализ и синтез. При поражении ядерной части рассеянные элементы могут до известной степени компенсировать выпавшую функцию ядра, что имеет огромное клиническое значение для восстановления данной функции.
До И. П. Павлова в коре различались двигательная зона, или двигательные центры, предцентральная извилина, и чувствительная зона, или чувствительные центры, расположенные позади s?lcus centr?lis. И. П. Павлов показал, что так называемая двигательная зона, соответствующая предцентральной извилине, есть, как и другие зоны мозговой коры, воспринимающая область (корковый конец двигательного анализатора). «Моторная область есть рецепторная область… Этим устанавливается единство всей коры полушарий» (И. П. Павлов). В настоящее время вся мозговая кора рассматривается как сплошная воспринимающая поверхность. Кора — это совокупность корковых концов анализаторов. С этой точки зрения мы и рассмотрим топографию корковых отделов анализаторов, т. е. главнейшие воспринимающие участки коры полушарий большого мозга.
Прежде всего рассмотрим корковые концы анализаторов, воспринимающих раздражения из внутренней среды организма (см. рис. 289, 299).
1. Ядро двигательного анализатора, т. е. анализатора проприоцептивных (кинестетических) раздражений, исходящих от костей, суставов, скелетных мышц и их сухожилий, находится в предцентральной извилине (поля 4 и 6) и l?bulus paracentr?lis. Здесь замыкаются двигательные условные рефлексы. Двигательные параличи, возникающие при поражении двигательной зоны, И. П. Павлов объясняет не повреждением двигательных эфферентных нейронов, а нарушением ядра двигательного анализатора, вследствие чего кора не воспринимает кинестетические раздражения и движения становятся невозможными. Клетки ядра двигательного анализатора заложены в средних слоях коры моторной зоны. В глубоких ее слоях (V, отчасти VI) лежат гигантские пирамидные клетки, представляющие собой эфферентные нейроны, которые И. П. Павлов рассматривает как вставочные нейроны, связывающие кору мозга с подкорковыми ядрами, ядрами черепных нервов и передними рогами спинного мозга, т. е. с двигательными нейронами. В предцентральной извилине тело человека, так же как и в задней, спроецировано вниз головой. При этом правая двигательная область связана с левой половиной тела и наоборот, ибо начинающиеся от нее пирамидные пути перекрещиваются частью в продолговатом, а частью в спинном мозге. Мышцы туловища, гортани, глотки находятся под влиянием обоих полушарий. Кроме предцентральной извилины, проприоцептивные импульсы (мышечно-суставная чувствительность) приходят и в кору постцентральной извилины.
2. Ядро двигательного анализатора, имеющего-отношение к сочетанному повороту головы и глаз в противоположную сторону, помещается в средней лобной извилине, в премоторной области (поле 8). Такой поворот происходит и при раздражении поля 17, расположенного в затылочной доле в соседстве с ядром зрительного анализатора. Так как при сокращении мышц глаза в кору мозга (двигательный анализатор, поле 8) всегда поступают не только импульсы от рецепторов этих мышц, но и импульсы от сетчатки (зрительный анализатор, поле 17), то различные зрительные раздражения всегда сочетаются с различным положением глаз, устанавливаемым сокращением мышц глазного яблока.
3. Ядро двигательного анализатора, посредством которого происходит синтез целенаправленных сложных профессиональных, трудовых и спортивных движений, помещается в левой (у правшей) нижней теменной дольке, в g?rus supramargin?lis (глубокие слои поля 40). Эти координированные движения, образованные по принципу временных связей и выработанные практикой индивидуальной жизни, осуществляются через связь g?rus supramargin?lis с предцентральной извилиной. При поражении поля 40 сохраняется способность к движению вообще, но появляется неспособность совершать целенаправленные движения, действовать — апраксия (праксия — действие, практика).
4. Ядро анализатора положения и движения головы — статический анализатор (вестибулярный аппарат) в коре мозга точно еще не локализован. Есть основания предполагать, что вестибулярный аппарат проецируется в той же области коры, что и улитка, т. е. в височной доле. Так, при поражении полей 21 и 20, лежащих в области средней и нижней височных извилин, наблюдается атаксия, т. е. расстройство равновесия, покачивание тела при стоянии. Этот анализатор, играющий решающую роль в прямохождении человека, имеет особенное значение для работы летчиков в условиях реактивной авиации, так как чувствительность вестибулярного аппарата на самолете значительно понижается.
5. Ядро анализатора импульсов, идущих от внутренностей и сосудов, находится в нижних отделах передней и задней центральных извилин. Центростремительные импульсы от внутренностей, сосудов, непроизвольной мускулатуры и желез кожи поступают в этот отдел коры, откуда исходят центробежные пути к подкорковым вегетативным центрам.
В премоторной области (поля 6 и 8) совершается объединение вегетативных и анимальных функций. Однако не следует считать, что только эта область коры влияет на деятельность внутренностей. На них оказывает влияние состояние всей коры полушарий большого мозга.
Нервные импульсы из внешней среды организма поступают в корковые концы анализаторов внешнего мира.
1. Ядро слухового анализатора лежит в средней части верхней височной извилины, на поверхности, обращенной к островку, — поля 41, 42, 52, где спроецирована улитка. Повреждение ведет к глухоте.
2. Ядро зрительного анализатора находится в затылочной доле — поля 17, 18, 19. На внутренней поверхности затылочной доли, по краям s?lcus calcar?nus, в поле 17 заканчивается зрительный путь. Здесь спроецирована сетчатка глаза, причем зрительный анализатор каждого полушария связан с полями зрения и соименными половинами сетчатки обоих глаз (например, левое полушарие связано с латеральной половиной левого глаза и медиальной правого). При поражении ядра зрительного анализатора наступает слепота. Выше поля 17 расположено поле 18, при поражении которого зрение сохраняется и только теряется зрительная память. Еще выше находится поле 19, при поражении которого утрачивается ориентация в непривычной обстановке.
3. Ядро обонятельного анализатора помещается в филогенетически древней части коры мозга, в пределах основания обонятельного мозга — ?ncus, отчасти гиппокампа (поле 11) (см. рис. 299, поля А и Е).
4. Ядро вкусового анализатора, по одним данным, находится в нижней части постцентральной извилины, близко к центрам мышц рта и языка, по другим — в ?ncus, в ближайшем соседстве с корковым концом обонятельного анализатора, чем объясняется тесная связь обонятельных и вкусовых ощущений. Установлено, что расстройство вкуса наступает при поражении поля 43.
Анализаторы обоняния, вкуса и слуха каждого полушария связаны с рецепторами соответствующих органов обеих сторон тела.
5. Ядро кожного анализатора (осязательная, болевая и температурная чувствительность) находится в постцентральной извилине (поля 1, 2, 3) и в коре верхней теменной области (поля 5 и 7). При этом тело спроецировано в постцентральной извилине вверх ногами, так что в верхней ее части расположена проекция рецепторов нижних конечностей, а в нижней — проекция рецепторов головы. Так как у животных рецепторы общей чувствительности особенно развиты на головном конце тела, в области рта, играющего огромную роль при захватывании пищи, то и у человека сохранилось сильное развитие рецепторов рта. В связи с этим область последних занимает в коре постцентральной извилины непомерно большую зону. Вместе с тем у человека в связи с развитием руки как органа труда резко увеличились рецепторы осязания в коже кисти, которая стала и органом осязания. Соответственно этому участки коры, соответствующие рецепторам верхней конечности, много больше таковых нижней конечности. Поэтому, если в постцентральную извилину врисовать фигуру человека головой вниз (к основанию черепа) и стопами вверх (к верхнему краю полушария), то надо нарисовать громадное лицо с несообразно большим ртом, большую руку, особенно кисть с большим пальцем, резко превосходящим остальные, небольшое туловище и маленькую ножку. Каждая постцентральная извилина связана с противоположной частью тела вследствие перекреста чувствительных проводников в спинном и частью в продолговатом мозге.
Частный вид кожной чувствительности — узнавание предметов на ощупь — стереогнозия (стереос — пространственный, гнозис — знание) связана с участком коры верхней теменной дольки (поле 7) перекрестно: левое полушарие соответствует правой руке, правое — левой руке. При поражении поверхностных слоев поля 7 утрачивается способность узнавать предметы на ощупь, при закрытых глазах.
Описанные корковые концы анализаторов расположены в определенных областях мозговой коры, которая, таким образом, представляет собой «грандиозную мозаику, грандиозную сигнализационную доску» (И. П. Павлов).
На эту «доску» благодаря анализаторам падают сигналы из внешней и внутренней среды организма. Эти сигналы, по И. П. Павлову, и составляют первую сигнальную систему действительности, проявляющуюся в форме конкретно-наглядного мышления (ощущения и комплексы ощущений — восприятия). Первая сигнальная система имеется и у животных. Но «в развивающемся животном мире на фазе человека произошла чрезвычайная прибавка к механизмам нервной деятельности. Для животного действительность сигнализируется почти исключительно раздражениями и следами их в полушариях большого мозга, непосредственно приходящими в специальные клетки зрительных, слуховых и других рецепторов организма. Это то, что и мы имеем в себе как впечатления, ощущения и представления от окружающей внешней среды, как общеприродной, так и от нашей социальной, исключая слово, слышимое и видимое. Это первая сигнальная система, общая у нас с животными. Но слово составило вторую, специально нашу сигнальную систему действительности, будучи сигналом первых сигналов…» (И. П. Павлов).
Таким образом, И. П. Павлов различает две корковые системы: первую и вторую сигнальные системы действительности, из которых сначала возникла первая сигнальная система (она имеется и у животных), а затем вторая — она имеется только у человека и является словесной системой. Вторая сигнальная система — это человеческое мышление, которое всегда словесно, ибо язык — это материальная оболочка мышления. Язык — это «…непосредственная действительность мысли» (Маркс К., Энгельс Ф. Соч., 2-е изд., т. 3, с. 448).
Путем весьма длительного повторения образовались временные связи между определенными сигналами (слышимые звуки и видимые знаки) и движениями губ, языка, мышц гортани, с одной стороны, и с реальными раздражителями или представлениями о них — с другой. Так, на базе первой сигнальной системы возникла вторая.
Отражая этот процесс филогенеза, у человека в онтогенезе сначала закладывается первая сигнальная система, а затем вторая. Чтобы вторая сигнальная система начала функционировать, требуется общение ребенка с другими людьми и приобретение навыков устной и письменной речи, на что уходит ряд лет. Если ребенок рождается глухим или теряет слух до того, как он начал говорить, то заложенная у него возможность устной речи не используется и ребенок остается немым, хотя звуки он произносить может. Точно так же, если человека не обучать чтению и письму, то он навсегда останется неграмотным. Все это свидетельствует о решающем влиянии окружающей среды для развития второй сигнальной системы. Последняя связана с деятельностью всей коры мозга, однако некоторые области ее играют особенную роль в осуществлении речи. Эти области коры являются ядрами анализаторов речи.
Поэтому для понимания анатомического субстрата второй сигнальной системы необходимо, кроме знания строения коры большого мозга в целом, учитывать также корковые концы анализаторов речи (рис. 300).
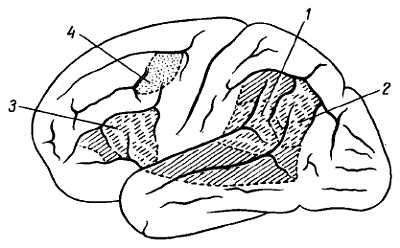
Рис. 300.
1 — слуховой (акустический) центр; 2 — зрительный (оптический) центр речи; 3 — двигательный центр устной речи; 4 — двигательный центр письменной речи.
1. Так как речь явилась средством общения людей в процессе их совместной трудовой деятельности, то двигательные анализаторы речи выработались в непосредственной близости от ядра общего двигательного анализатора.
Двигательный анализатор артикуляции речи (речедвигательный анализатор) находится в задней части нижней лобной извилины (поле 44), в непосредственной близости от нижнего отдела моторной зоны. В нем происходит анализ раздражений, проходящих от мускулатуры, участвующей в создании устной речи. Эта функция сопряжена с двигательным анализатором мышц губ, языка и гортани, находящимся в нижнем отделе предцентральной извилины, чем и объясняется близость речедвигательного анализатора к двигательному анализатору названных мышц. При поражении поля 44 сохраняется способность производить простейшие движения речевой мускулатуры, кричать и даже петь, но утрачивается возможность произносить слова — двигательная афазия (фазис — речь). Впереди поля 44 расположено поле 45, имеющее отношение к речи и пению. При поражении его возникает вокальная амузия — неспособность петь, составлять музыкальные фразы, а также аграмматизм — неспособность составлять из слов предложения.
2. Так как развитие устной речи связано с органом слуха, то в непосредственной близости к звуковому анализатору выработался слуховой анализатор устной речи. Его ядро помещается в задней части верхней височной извилины, в глубине латеральной борозды (поле 42). Благодаря слуховому анализатору различные сочетания звуков воспринимаются человеком как слова, которые означают различные предметы и явления и становятся сигналами их (вторыми сигналами). С помощью его человек контролирует свою, речь и понимает чужую. При поражении его сохраняется способность слышать звуки, но теряется способность понимать слова — словесная глухота, или сенсорная афазия. При поражении поля 22 (средняя треть верхней височной извилины) наступает музыкальная глухота: больной не знает мотивов, а музыкальные звуки воспринимаются им как беспорядочный шум.
3. На более высокой ступени развития человечество научилось не только говорить, но и писать. Письменная речь требует определенных движений руки при начертании букв или других знаков, что связано с двигательным анализатором (общим). Поэтому двигательный анализатор письменной речи помещается в заднем отделе средней лобной извилины, вблизи зоны предцентральной извилины (моторная зона). Деятельность этого анализатора связана с анализатором необходимых при письме заученных движений руки (поле 40 в нижней теменной дольке). При повреждении поля 40 сохраняются все виды движения, но теряется способность тонких движений, необходимых для начертания букв, слов и других знаков (аграфия).
4. Так как развитие письменной речи связано и с органом зрения, то в непосредственной близости к зрительному анализатору выработался зрительный анализатор письменной речи, который, естественно, расположен вблизи s?lcus calcar?nus, в g?rus angul?ris (поле 39). При повреждении нижней теменной дольки сохраняется зрение, но теряется способность читать (алексия), т. е. анализировать написанные буквы и слагать из них слова и фразы.
Все речевые анализаторы закладываются в обоих полушариях, но развиваются только с одной стороны (у правшей — слева, у левшей — справа) и функционально оказываются асимметричными. Эта связь между двигательным анализатором руки (органа труда) и речевыми анализаторами объясняется тесной связью между трудом и речью, оказавшими решающее влияние на развитие мозга.
«…Труд, а затем и вместе с ним членораздельная речь…» привели к развитию мозга (Маркс К., Энгельс Ф. Соч., 2-е изд., т. 20, с. 490). Этой связью пользуются и в лечебных целях. При поражении речедвигательного анализатора сохраняется элементарная двигательная способность речевых мышц, но утрачивается возможность устной речи (моторная афазия). В этих случаях иногда удается восстановить речь длительным упражнением левой руки (у правшей), работа которой благоприятствует развитию зачаточного правостороннего ядра речедвигательного анализатора.
Анализаторы устной и письменной речи воспринимают словесные сигналы (как говорил И. П. Павлов, сигналы сигналов, или вторые сигналы), что составляет вторую сигнальную систему действительности, проявляющуюся в форме абстрактного отвлеченного мышления (общие представления, понятия, умозаключения, обобщения), которое свойственно только человеку. Однако морфологическую основу второй сигнальной системы составляют не только указанные анализаторы. Так как функция речи является филогенетически наиболее молодой, то она и наименее локализована. Так как кора растет по периферии, то наиболее поверхностные слои коры имеют отношение ко второй сигнальной системе. Эти слои состоят из большого числа нервных клеток (15 млрд) с короткими отростками, благодаря которым создается возможность неограниченной замыкательной функции, широких ассоциаций, что и составляет сущность деятельности второй сигнальной системы. При этом вторая сигнальная система функционирует не отдельно от первой, а в тесной связи с ней, точнее на основе ее, так как вторые сигналы могут возникнуть лишь при наличии первых. «Основные законы, установленные в работе первой сигнальной системы, должны также управлять и второй, потому что это работа все той же нервной ткани» (И. П. Павлов).
Учение И. П. Павлова о двух сигнальных системах дает материалистическое объяснение психической деятельности человека и составляет естественнонаучную основу теории отражения В. И. Ленина. Согласно этой теории, в нашем сознании в форме субъективных образов отражается объективный реальный мир, существующий независимо от нашего сознания.
Ощущение — это субъективный образ объективного мира. «…Ощущение… есть превращение энергии внешнего раздражения в факт сознания» (Ленин В. И. Полн. собр. соч., т. 18, с. 46).
В рецепторе внешнее раздражение, например световая энергия, превращается в нервный процесс, который в коре мозга становится ощущением.
Одно и то же количество и качество энергии, в данном случае световой, у здоровых людей вызовет в коре мозга ощущение зеленого цвета (субъективный образ), а у больного дальтонизмом (благодаря иному строению сетчатки глаза) — ощущение красного цвета.
Следовательно, световая энергия — это объективная реальность, а цвет — субъективный образ, отражение ее в нашем сознании, зависящее от устройства органа чувств (глаза).
Значит, с точки зрения ленинской теории отражения мозг может быть охарактеризован как орган отражения действительности.
После всего сказанного о строении центральной нервной системы можно отметить «человеческие» признаки строения мозга, т. е. специфические черты строения его, отличающие человека от животных (рис. 301, 302).
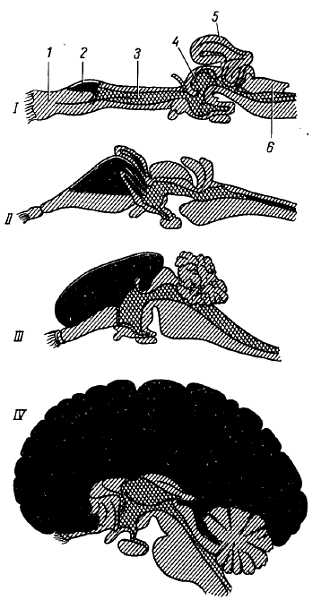
Рис. 301.
I — акула: II — ящерица; III — кролик; IV — человек; 1 — lobus olfactorius; 2 — corpus striatum; 3 — diencephaIon; 4 — mesencephalon; 5 — cerebellum; 6 — medulla oblongata.
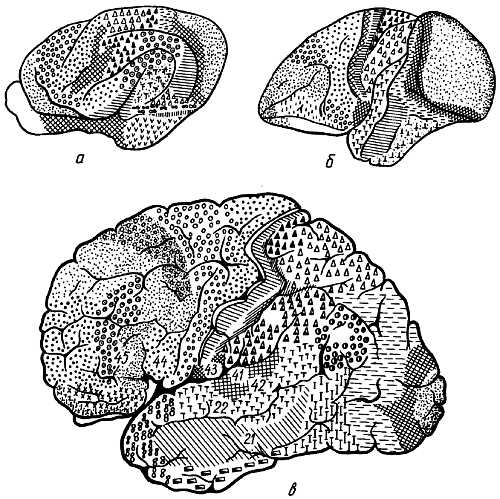
Рис. 302
1. Преобладание головного мозга над спинным. Так, у хищных (например, у кошки) головной мозг в 4 раза тяжелее спинного, у приматов (например, у макак) — в 8 раз, а у человека — в 45 раз (масса спинного мозга 30 г, головного — 1500 г). Спинной мозг составляет у млекопитающих 22–48 % массы головного мозга, у гориллы — 5–6 %, у человека — только 2 %.
2. Масса мозга. По абсолютной массе мозга человек не занимает первого места, так как у крупных животных мозг тяжелее, чем у человека (1500 г): у дельфина — 1800 г, у слона — 5200 г, у кита — 7000 г. Чтобы вскрыть истинные отношения массы мозга к массе тела, используют так называемый квадратный указатель мозга, т. е. произведение абсолютной массы мозга на относительную. Этот указатель позволил выделить человека из всего животного мира. Так, у грызунов он равен 0,19, у хищных — 1,14, у китообразных (дельфин) — 6,27, у человекообразных обезьян — 7,35, у слонов — 9,82 и, наконец, у человека — 32,0.
3. Преобладание плаща над мозговым стволом, т. е. нового мозга (neenc?phalon) над древним (paleenc?phalon).
4. Наивысшее развитие лобной доли большого мозга. На лобные доли приходится у низших обезьян 8-12 % всей поверхности полушарий, у антропоидных обезьян — 16 %, у человека — 30 %.
5. Преобладание новой коры полушарий большого мозга над старой.
6. Преобладание коры над подкоркой, которое у человека достигает максимальных цифр: кора составляет 53,7 % всего объема мозга, а базальные ядра — только 3,7 %
7. Борозды и извилины увеличивают площадь коры серого вещества, поэтому чем больше развита кора полушарий большого мозга, тем больше и складчатость мозга. Увеличение складчатости достигается большим развитием мелких борозд третьей категории, глубиной борозд и их асимметричным расположением. Ни у одного животного нет одновременно такого большого числа борозд и извилин, при этом столь глубоких и асимметричных, как у человека.
8. Наличие второй сигнальной системы, анатомическим субстратом которой являются самые поверхностные слои мозговой коры.
Подводя итоги изложенному, можно сказать, что специфическими чертами строения мозга человека, отличающими его от мозга самых высокоразвитых животных, является максимальное преобладание молодых частей центральной нервной системы над старыми: головного мозга над спинным, плаща над стволом, новой коры над старой, поверхностных слоев мозговой коры над глубокими.
Ложность «теории» расизма в учении о мозге
Чтобы оправдать стремление империалистических кругов к мировому господству, находящиеся на службе эксплуататорских классов реакционные ученые создали «теорию» расизма, согласно которой народы мира изначально делятся на передовые и отсталые, а человеческие расы — на высшие и низшие. Высшие расы имеют, по мнению расистов, право на покорение низших не только в силу экономической и политической отсталости последних, но и вследствие якобы их более низкой биологической организации.
В качестве аргументов для отнесения к более низкой организации привлекаются некоторые признаки строения мозга, а именно: сравнительно меньшие масса и объем мозга, меньшее число борозд и извилин, редкие вариации их, наличие борозд, более выраженных у приматов, например обезьянья борозда в затылочной доле, а также ряд других признаков.
Однако все эти особенности строения мозга не могут служить признаками низшего развития. В самом деле, если взять абсолютную массу мозга, то она не может являться показателем умственного развития человека, так как у гениальных людей можно встретить мозг самой различной массы. Масса мозга людей колеблется от 1100 до 2000 г. Тяжелый мозг встречается не только у умственно одаренных людей, но и у людей среднего развития, а также у эпилептиков и идиотов. Так, самый тяжелый мозг из всех известных до настоящего времени (2850 г) принадлежал идиоту-эпилептику 21 года. Приводимая А. Якобом сводка цифр массы мозга 50 выдающихся деятелей различных специальностей показывает, что масса их мозга колеблется в широких границах, так что никоим образом нельзя провести прямые параллели между массой мозга и одаренностью. Так, если сравнить массу мозга крупнейших писателей — И. С. Тургенева и Анатоля Франса, то при одинаковом характере их одаренности мозг И. С. Тургенева был более тяжелым (2012 г), а мозг Анатоля Франса вдвое легче (1017 г), что не помешало Анатолю Франсу проявить свой талант. То же наблюдается и при сравнении мозга других выдающихся людей, например поэтов Байрона (2238 г) и Уитмена (1282 г), ученых-зоологов Кювье (1830 г) и Агассица (1495 г) и др.
Такая же картина наблюдается при сравнении у различных гениальных людей абсолютного объема мозга и черепа: например, у Гете окружность головы 60 см, у Данте — 54 см, что не помешало Данте написать свое бессмертное произведение «Божественная комедия».
Как показали исследования Л. Я. Пинеса, обезьянья борозда с одинаковой частотой встречается на внутренней поверхности затылочной доли мозга у представителей различных рас и у интеллектуально выдающихся лиц. Развитие других борозд и извилин также подвержено различным вариациям, с одинаковой частотой встречающихся у разных народов. Ряд объективных буржуазных исследователей высказываются, что на основании наличия различных борозд нельзя делать заключение об умственной одаренности. Таким образом, отмеченные особенности строения мозга являются не расовыми признаками, а вариантами индивидуальной изменчивости, которой подвержены все органы, в том числе и мозг. Закономерное нарастание массы и объема мозга действительно имеет место в эволюции человека, но оно происходит в течение сотен тысячелетий. Так, у человекообразных обезьян масса мозга 400–500 г, а у современного человека — 1100–2000 г (в среднем 1500 г).
Что же касается современных людей, то колебания массы и объема мозга не отражают степени умственного развития. Культурная и политическая отсталость народов обусловливается не биологической организацией (строение мозга и всего тела человека), а социальными условиями жизни общества. Яркий пример этого мы видим в Индии и других бывших колониальных странах. Там раньше, чем в Европе, возникла древняя и весьма высокая культура, создавшая замечательные памятники искусства, зодчества и литературы. Однако после порабощения Индии англичанами, за три столетия колониального гнета, развитие индийского народа резко затормозилось, и он отстал от завоевавших его европейцев. Теперь, когда порабощенные народы сбросили ярмо колониализма и снова стали свободными, они быстро идут по пути социального прогресса. Таким образом, необходимо видеть реакционную политическую сущность расизма и разоблачать ее, опираясь на строго научные морфологические факты.
Оболочки головного мозга
Оболочки головного мозга, meninges, составляют непосредственное продолжение оболочек спинного мозга — твердой, паутинной и мягкой.
Твердая оболочка, d?ra m?ter enc?phali, — плотная белесоватая соединительнотканная оболочка, лежащая снаружи от остальных оболочек. Наружная ее поверхность непосредственно прилежит к черепным костям, для которых твердая оболочка служит надкостницей, в чем состоит ее отличие от такой же оболочки спинного мозга. Внутренняя поверхность, обращенная к мозгу, покрыта эндотелием и вследствие этого гладкая и блестящая. Между ней и паутинной оболочкой мозга находится узкое щелевидное пространство, sp?tium subdur?le, заполненное небольшим количеством жидкости. Местами твердая оболочка расщепляется на два листка. Такое расщепление имеет место в области венозных синусов (см. ниже), а также в области ямки у верхушки пирамиды височной кости (impressio trigemini), где лежит узел тройничного нерва. Твердая оболочка отдает со своей внутренней стороны несколько отростков, которые, проникая между частями мозга, отделяют их друг от друга (рис. 303).
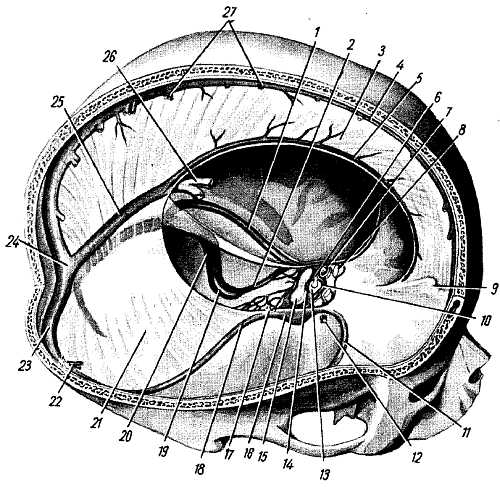
Рис. 303
1, 18 — sinus petrosus superior (dexter et sinister); 2 — sinus petrosus inferior; 5 — falx cerebri; 4 — sinus sagittalis superior; 5 — sinus sagittalis inferior; 6 — infundibulum; 7 — a. carotis interna; 8 — n. opticus; 9 — crista galli; 10, 14 — sinus intercavernosus: 11 — sinus sphenoparietalis; 12 — cerebri media; 13 — diaphragma sellae; 15 — dorsum sellae; 16 — sinus cavernosus; 17 — plexus basillaris; 19 — bulbus superior v. jugular is internae; 20 — sinus sigmoideus; 21 — tentorium cerebelli; 22 — vv. cerebri inferiores; 23 — sinus transversus; 24 — confluens sinuum; 25 — sinus rectus; 26 — v. cerebri magna; 27 — vv. cerebri superiores.
Falx c?rebri, серп большого мозга, расположен в сагиттальном направлении между обоими полушариями большого мозга. Прикрепляясь по средней линии черепного свода к краям s?lcus s?nus sagitt?lis superi?ris, он своим передним узким концом прирастает к cr?sta g?lli, а задним широким срастается с верхней поверхностью мозжечкового намета.
Tent?rium cereb?lli, намет мозжечка, представляет горизонтально натянутую пластинку, слегка выпуклую кверху наподобие двускатной крыши. Пластинка эта прикрепляется по краям s?lcus s?nus transv?rsi затылочной кости и вдоль верхней грани пирамиды височной кости на обеих сторонах до proc?ssus clino?deus posterior клиновидной кости. Намет мозжечка отделяет затылочные доли большого мозга от нижележащего мозжечка.
Falx cereb?lli, серп мозжечка, располагается, так же как и серп большого мозга, по средней линии вдоль cr?sta occipit?lis int?rna до большого отверстия затылочной кости, охватывая последнее по бокам двумя ножками; этот невысокий отросток вдается в заднюю вырезку мозжечка.
Diaphr?gma s?llae, диафрагма седла, пластинка, ограничивающая сверху вместилище для гипофиза на дне турецкого седла. В середине она прободается отверстием для пропуска воронки, infund?bulum, к которой прикрепляется hyp?physis.
Кровеносные сосуды твердой оболочки питают также кости черепа и образуют на внутренней пластинке последних вдавления, s?lci men?ngei. Из артерий самая крупная a. men?ngea m?dia, ветвь a. maxill?ris, проходящая в череп через for?men spin?sum клиновидной кости. В передней черепной ямке разветвляется небольшая ветвь из a. ophth?lmica, а в задней — веточки из a. phar?ngea asc?ndens, из a. vertebr?lis и из a. occipit?lis, проникающие через for?men masto?deum. Вены твердой оболочки сопровождают соответствующие артерии, обычно по две, и впадают частью в синусы, частью в pl?xus pterygo?deus.
Нервы. Твердая оболочка иннервируется тройничным нервом.
Кроме собственных вен, твердая оболочка содержит ряд вместилищ, собирающих кровь из мозга и называемых синусами твердой оболочки, s?nus d?rae m?tris (см. рис. 303).
Синусы представляют венозные, лишенные клапанов каналы (треугольные в поперечном сечении), залегающие в толще самой твердой оболочки по местам прикрепления ее отростков к черепу и отличающиеся от вен строением своих стенок. Последние образованы туго натянутыми листками твердой оболочки, вследствие чего не спадаются при разрезе и при ранении зияют. Неподатливость стенок венозных синусов обеспечивает свободный отток венозной крови при смене внутричерепного давления, что важно для бесперебойной деятельности головного мозга, чем и объясняется наличие таких венозных синусов только в черепе.
Имеются следующие синусы:
S?nus transv?rsus — самый большой и широкий, расположен по заднему краю tent?rium cereb?lli в s?lcus s?nus transv?rsi затылочной кости, откуда спускается как s?nus sigmo?deus в s?lcus sinus sigmo?dei и далее у for?men jugul?re переходит в устье v. jugul?ris int?rna. Благодаря этому поперечный синус с сигмовидным служит главным коллектором для всей венозной крови черепной полости. В него частью непосредственно, частью опосредованно впадают все остальные синусы. Непосредственно в него впадают:
S?nus sagitt?lis sup?rior идет по верхнему краю falx c?rebri вдоль всего s?lcus sinus sagitt?lis superi?ris or cr?sta g?lli до protuber?ntia occipit?lis int?rna; по бокам s?nus sagitt?lis sup?rior, в толще твердой оболочки, заложены так называемые кровяные озера — небольшие полости, сообщающиеся с одной стороны с синусом и диплоическими венами, а с другой — с венами твердой оболочки и мозга.
S?nus occipit?lis — как бы продолжение предыдущего вдоль места прикрепления falx cereb?lli к cr?sta occipit?lis int?rna и далее (после раздвоения) по обоим краям for?men m?gnum затылочной кости.
S?nus r?ctus на линии прикрепления falx c?rebri к tent?rium cereb?lli. Он принимает спереди s?nus sagitt?lis inf?rior, идущий вдоль нижнего свободного края falx c?rebri, а также v. c?rebri m?gna, по которой кровь оттекает из глубоких частей мозга.
В месте, где сходятся названные синусы (s?nus transv?rsus, s?nus sagitt?lis sup?rior, s?nus r?ctus и s?nus occipit?lis), образуется общее расширение, известное под именем стока синусов, c?nfluens s?nuum. На основании черепа сбоку турецкого седла расположен пещеристый синус, s?nus cavern?sus, имеющий вид или венозного сплетения, или широкой лакуны, окружающей внутреннюю сонную артерию. Он соединяется с таким же синусом другой стороны двумя поперечными анастомозами, s?nus intercavern?si, проходящими спереди и сзади f?ssa hypophysi?lis, вследствие чего в области турецкого седла образуется венозное кольцо.
Пещеристый синус представляет сложный анатомический комплекс, в состав которого, кроме самого синуса, входят внутренняя сонная артерия, нервные стволы и окружающая их соединительная ткань. Все эти образования составляют как бы особый прибор, играющий важную роль в регуляции внутричерепного тока венозной крови. Спереди в пещеристый синус вливаются v. ophth?lmica sup?rior, проходящая через верхнюю глазничную щель, а также нижний конец s?nus sphenopariet?lis, идущего вдоль края ?lae min?ris.
Отток крови из s?nus cavern?sus совершается в два лежащих сзади синуса: s?nus petr?sus sup?rior et inf?rior, заложенные в соименных желобках, s?lcus s?nus petr?si superi?ris et inferi?ris. Оба s?nus petr?si inferi?res соединяются между собой несколькими венозными каналами, которые лежат в толще твердой оболочки на базилярной части затылочной кости и называются в своей совокупности pl?xus basil?ris. Pl?xus basil?ris сообщается с венозными сплетениями позвоночного канала, через которые таким образом оттекает кровь из полости черепа.
Главным путем оттока крови из синусов служат внутренние яремные вены, но, кроме того, венозные синусы соединяются с венами наружной поверхности черепа посредством так называемых эмиссарных вен, vv. emiss?riae, проходящих через отверстия в черепных костях (for?men parietale, for?men masto?deum, can?lis condyl?ris см. «Остеология»). Такую же роль играют небольшие вены, выходящие из черепа вместе с нервами через for?men ov?le, for?men rot?ndum и can?lis hypoglossalis. В синусы твердой оболочки также впадают v?nae dipl?icae, вены губчатого вещества костей черепа; другим концом они могут иметь связь с наружными венами головы. V?nae dipl?icae представляют анастомозирующие друг с другом каналы, выстланные изнутри слоем эндотелия и проходящие в губчатом веществе плоских костей черепа.
Паутинная оболочка, arachno?dea enc?phali (рис. 304), так же как и в спинном мозге, отделяется от твердой оболочки капиллярной щелью субдурального пространства. Паутинная оболочка не заходит в глубину борозд и углублений мозга, как p?a m?ter, но перекидывается через них в виде мостиков, вследствие чего между ней и мягкой оболочкой находится подпаутинное пространство, c?vitas subarachnoide?lis, которое наполнено прозрачной жидкостью.
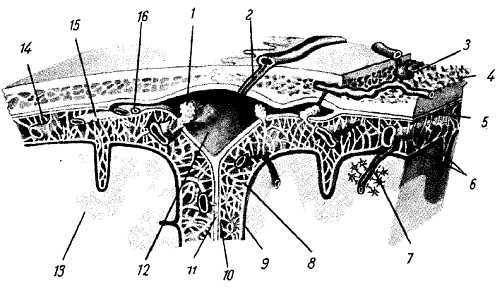
Рис. 304
1, 16 — granulationes arachnojdeales; 2 — v. emissaria; 3 — v. diploica; 4 — diplo?; 5 — dura mater encephali; 6 — trabeculae arachnoideales; 7 — spatium perjvasculare; 8 — cavitas subarachnoidealis; 9 — pia mater; 10 — arachnoidea; 11 — falx cerebri; 12 — sinus sagittalis superior; 13 — cortex cerebri; 14 — r. corticalis a. cerebri; 15 — r. corticalis v. cerebri.
В некоторых местах, преимущественно на основании мозга, подпаутинные пространства развиты особенно сильно, образуя широкие и глубокие вместилища спинномозговой жидкости, называемые цистернами (рис. 305).
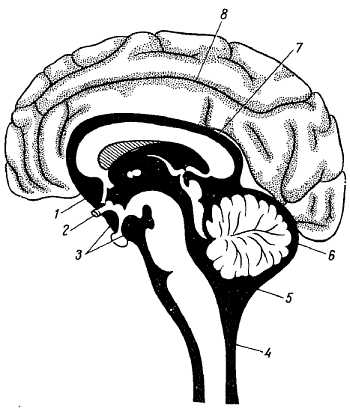
Рис. 305
1 — cisterna chiasmatis; 2 — chiasma opticum; 3 — cisterna interpeduncularis; 4 — подпаутинное пространство спинного мозга; 5 — cisterna сеrebellomedullaris; 6 — arachnoidea; 7 — подпаутинное пространство над мозолистым телом; 8 — подпаутинное пространство в бороздах.
Имеются следующие цистерны:
1. Cist?rna cerebellomedull?ris (самая большая) между задним краем мозжечка и продолговатым мозгом.
2. Cisierna interpeduncul?ris между ped?nculi cerebri.
3. Cisterna chiasmatis впереди chiasma ?pticum.
4. Cist?rna f?ssae later?lis c?rebri в соименной ямке.
Все подпаутинные пространства широко сообщаются между собой и у большого отверстия затылочной кости непосредственно продолжаются в подпаутинное пространство спинного мозга. Кроме того, они находятся в прямом сообщении с желудочками мозга через отверстия в области задней стенки IV желудочка: apert?ra medi?na ventr?culi qu?rti, открывающееся в cist?rna cerebellomedull?ris, и apert?ra later?lis ventr?culi IV. В подпаутинных пространствах залегают мозговые сосуды, которые соединительнотканными перекладинами, trab?culae arachnoide?les, и окружающей жидкостью предохраняются от сдавления.
Особенностью строения паутинной оболочки являются так называемые грануляции паутинной оболочки, granulati?nes arachnoide?les, представляющие выросты паутинной оболочки в виде кругловатых телец серо-розового цвета, вдающихся в полость венозных синусов или же в лежащие рядом кровяные озера (см. рис. 304). Они имеются у детей и у взрослых, но наибольшей величины и многочисленности достигают в старости. Увеличиваясь в размерах, грануляции своим давлением на черепные кости образуют на внутренней поверхности последних углубления, известные в остеологии под названием fov?olae granul?res. Грануляции служат для оттока спинномозговой жидкости в кровяное русло путем фильтрации.
Мягкая оболочка, p?a m?ter enc?phali, тесно прилегает к мозгу, заходя во все борозды и щели его поверхности, и содержит кровеносные сосуды и сосудистые сплетения (см. рис. 287). Между оболочкой и сосудами существует пери васкулярная щель, сообщающаяся с подпаутинным пространством.
Спинномозговая жидкость
Спинномозговая жидкость, l?quor cerebrospin?lis, наполняющая подпаутинные пространства головного и спинного мозга и мозговые желудочки, резко отличается от других жидкостей организма. С ней сходны только эндо- и перилимфа внутреннего уха и водянистая влага глаза. Выделение спинномозговой жидкости происходит путем секреции из pl?xus choro?dei, эпителиальная обкладка которых имеет характер железистого эпителия. Аппарат, продуцирующий l?quor cerebrospin?lis, обладает свойством пропускать в жидкость одни вещества и задерживать другие (гематоэнцефалический барьер), что имеет большое значение для предохранения мозга от вредных влияний. Таким образом, по своим особенностям спинномозговая жидкость является не только механическим защитным приспособлением для мозга и лежащих на его основании сосудов, но и специальной внутренней средой, которая необходима для правильного функционирования центральных органов нервной системы. Пространство, в котором помещается l?quor cerebrospin?lis, замкнуто. Отток жидкости из него совершается путем фильтрации главным образом в венозную систему через посредство грануляций паутинной оболочки, а отчасти также и в лимфатическую систему через влагалища нервов, в которые продолжаются мозговые оболочки.
Сосуды головного мозга
Артерии большого мозга происходят из ветвей a. car?tis int?rna и a. basil?ris, образующих на основании мозга circulus arteri?sus c?rebri (см. «Сосудистая система»). На поверхности каждого полушария разветвляются передняя, средняя и задняя мозговые артерии. A. c?rebri ant?rior снабжает кровью медиальную поверхность полушария до s?lcus parietooccipit?lis, на наружной его поверхности верхнюю лобную извилину и верхний край теменной доли, а на нижней поверхности полушария — g?rus r?ctus лобной доли. A. c?rebri m?dia снабжает кровью островок, обе центральные извилины, нижнюю лобную извилину и большую часть средней лобной извилины, теменную долю и верхнюю и среднюю височные извилины. A. c?rebri post?rior разветвляется на медиальной, нижней и латеральной поверхностях височной и затылочной долей, за исключением верхней и средней височной извилин (рис. 306).
Рис. 306
а — наружная поверхность: зоны a. cerebri media (светлые); зоны аа. cerebri anterior et posterior (заштрихованы); б — внутренняя поверхность: зоны a. cerebri anterior (мелкие точки), media (клетки) и posterior (линии). Мозолистое тело (красный цвет).
Перечисленные артерии своими разветвлениями в p?a m?ter образуют артериальную сеть, из которой проникают отвесно в толщу мозгового вещества: 1) кортикальные артерии — маленькие веточки, разветвляющиеся только в мозговой коре, и 2) медуллярные артерии, которые, пройдя кору, идут в белое вещество.
Со стороны основания мозга входят центральные артерии.
Кортикальные, медуллярные и центральные артерии анастомозируют друг с другом, образуя единую сосудистую сеть. Мозжечок получает кровь из трех артерий с каждой стороны. Две a. cereb?lli inf?rior anterior (ветвь a. basil?ris) и a. cereb?lli inf?rior post?rior (ветвь a. vertebr?lis), разветвляются на нижней поверхности мозжечка, третья же ветвь, a. cereb?lli sup?rior (ветвь a. basil?ris), идет на его верхнюю поверхность. От a. cereb?lli sup?rior снабжаются также нижние холмики крыши среднего мозга, а верхние холмики получают свои веточки от a. c?rebri post?rior. Артерии остальных частей головного мозга, относящиеся к мосту и продолговатому мозгу, происходят от a. vertebr?lis, a. basil?ris и их ветвей.
Кроме описанных артериальных сосудов, имеются еще особые артерии сосудистых сплетений в числе четырех на каждой стороне.
Вены большого мозга разделяются на поверхностные и глубокие. Поверхностные вены большей частью собирают кровь из мозговой коры и вливаются частью в s?nus sagitt?lis sup?rior (верхние вены), частью (нижние вены) в s?nus transv?rsus и синусы основания черепа. Вены лишены клапанов и отличаются своими многочисленными соустьями. Глубокие вены собирают кровь из центральных серых ядер и желудочков мозга и сливаются в одну большую v. c?rebri m?gna, впадающую в s?nus r?ctus.
Вены мозжечка составляют группы: верхние изливают кровь в s?nus r?ctus и v. c?rebri m?gna, нижние — в s?nus transv?rsus, sigmo?deus, petr?sus inf?rior.
Рентгенологическое исследование центральной нервной системы (рис. 307) позволяет видеть мозг живого человека без вскрытия черепа на светлом фоне нейтрального газа, введенного в подпаутинное пространство. Такой метод называется энцефалографией.
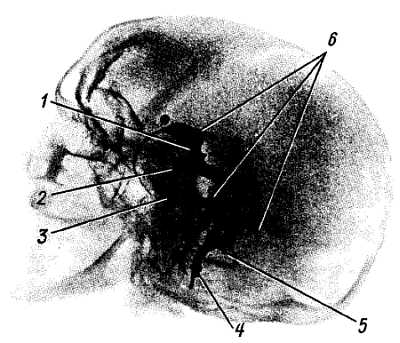
Рис. 307
1 — for. interventriculare; 2 — infundibulum; 3, 4 — подпаутинное пространство; 5 — ventriculus quartus; 6 — ventriculus lateralis: cornu anterius, cornu inferius, cornu posterius.
На энцефалограммах видны контуры головного мозга и его отдельных частей и выявляется ветвистый рисунок мозговых борозд. Введенный в подпаутинное пространство газ или контрастная жидкость попадают в систему мозговых камер, давая изображение мозговых желудочков (вентрикулография).
На боковой вентрикулограмме ясно видны: передний рог, центральная часть, задний и нижний рога бокового желудочка; III и IV желудочки на боковых снимках видны не всегда. Увеличение их является признаком патологического расширения.
На снимках, сделанных в затылочном положении (к пленке прилегает затылочная область; ход лучей сагиттальный), видна характерная симметричная фигура боковых желудочков, напоминающая бабочку. Правую и левую половины фигуры бабочки (как бы крылья ее) разделяет вертикально идущая линия — s?ptum pell?cidum, отделяющая оба боковых желудочка. Книзу от нее располагается узкая светлая щель — III желудочек. Ниже последнего в единичных случаях замечается узкая щелевидная тень IV желудочка. При патологическом расширении мозговых желудочков фигура бабочки искажается, а III и IV желудочки увеличиваются и становятся ясно заметными.
С помощью новейшего метода рентгенологического исследования — компьютерной (вычислительной) томографии можно получать рентгеновское изображение любого слоя головного мозга в любой плоскости; в частности удается видеть все части мозговых желудочков без контрастирования.


![Дочки-матери [Все, о чем вам не рассказывала ваша мама и чему стоит научить свою дочь]](https://img-lib.med-tutorial.ru/3386939452/cover.jpg)



