СХЕМА ПРОВОДЯЩИХ ПУТЕЙ НЕРВНОЙ СИСТЕМЫ
Проводящие пути с точки зрения направления проведения импульса могут быть разделены на две большие группы — афферентные и эфферентные. Афферентные проводящие пути составляют среднее звено — кондуктор того или иного анализатора; поэтому часть их будет рассмотрена вместе с соответствующими анализаторами (см. «Органы чувств»).
АФФЕРЕНТНЫЕ (ВОСХОДЯЩИЕ) ПРОВОДЯЩИЕ ПУТИ
Поскольку организм получает раздражение как из внешней среды, так и из внутренней, имеются пути, несущие импульсы от рецепторов, воспринимающих внешние раздражения, и от рецепторов, воспринимающих внутренние раздражения.
Проводящие пути от рецепторов внешних раздражений
Рецепторы, воспринимающие внешние раздражения, называются экстероцепторами. На ранних стадиях эволюции они были заложены главным образом в наружных покровах тела, что необходимо для восприятия внешних раздражений, почему и у человека они развиваются в эмбриогенезе из наружного зародышевого листка — эктодермы. Исключение представляет орган вкуса, тесно связанный функционально с пищеварительной системой и поэтому развивающийся из энтодермы (эпителия глоточных карманов). В дальнейшем с усложнением организации животных и усложнением их образа жизни те из экстероцепторов, которые имели жизненно важное значение, начали усиленно развиваться и усложняться в своей организации, приобретая строение особых органов, воспринимающих раздражения, источники которых находятся на известном расстоянии от организма и потому называемые дистантными. Это — рецепторы слуха, зрения и обоняния. Остальные рецепторы наружных покровов остались заложенными в коже, составляя периферическую часть кожного анализатора. Проводящие пути от рецепторов звука, света, вкуса и обоняния будут рассмотрены при описании соответствующих анализаторов в разделе эстезиологии. Здесь будут изложены проводящие пути кожного анализатора.
Проводящие пути кожного анализатора
Афферентные волокна кожного анализатора несут в кору большого мозга тактильные раздражения, чувство стереогноза (узнавание предмета на ощупь), болевые и температурные раздражения. В связи с этим их можно разбить на несколько групп.
Проводящие пути тактильной чувствительностикожи (чувство осязания) (рис. 348[рисунка нет]). Tractus gangliospinothalamocortic?lis. Рецептор находится толще кожи. Кондуктор состоит из 3 нейронов. Клеточное тело первого нейрона помещается в спинномозговом узле, который представляет собой скопление клеток периферических нейронов всех видов чувствительности. Отходящий от клеток этого узла отросток делится на две ветви, из которых периферическая идет в составе кожного нерва от рецептора, а центральная в составе заднего корешка идет в задние канатики спинного мозга, где в свою очередь делится на восходящую и нисходящую ветви. Концевые разветвления и коллатерали одной части волокон заканчиваются в задних рогах спинного мозга в subst?ntia gelatin?sa (эта часть тракта носит название tr?ctus gangliospin?lis), другая часть восходящих волокон не заходит в задние pora, а идет в задних канатиках спинного мозга и достигает в составе fasc?culus gr?cilis et cune?tus соименных ядер продолговатого мозга, n?cleus gr?cilis и n?cleus cune?tus (эта часть тракта называется tractus gangliobulb?ris).
В задних рогах спинного мозга и в названных ядрах продолговатого мозга помещается клеточное тело второго нейрона. Аксоны клеток, заложенных в задних рогах, пересекают срединную плоскость в commiss?ra ?lba и входят в состав расположенного в боковом канатике противоположной стороны tr?ctus spinoth?lamicus later?lis, который они и образуют (см. рис. 270).
Важно иметь в виду, что перекрест волокон спинно-таламических пучков происходит не на уровне вступления соответствующего заднего корешка в спинной мозг, а на 2–3 сегмента выше. Этот факт имеет существенное значение для клиники, так как при одностороннем повреждении этого пучка расстройство кожной чувствительности на противоположной стороне наблюдается не на уровне поражения, а книзу от него.
Этот пучок через стволовую часть головного мозга достигает таламуса. По пути он устанавливает связь с двигательными ядрами мозгового ствола и черепных нервов, по которым возникают головные рефлексы при раздражении кожи, например движение глаз при раздражении кожи руки. Аксоны клеток второго звена, заложенных в ядрах продолговатого мозга, также достигают таламуса по тракту, носящему название lemn?scus medi?lis, который в продолговатом мозге переходит на противоположную сторону, образуя перекрест медиальной петли (decuss?tio lemnisc?rum) (рис. 349).
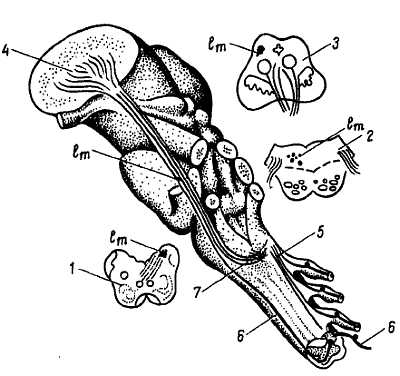
Рис. 349
lm — lemniscus medialis; 1, 2, 3 — поперечные сечения продолговатого мозга, моста и среднего мозга с обозначением положения медиальной петли (lm) в толще этих образований; 4 — nucl. lateralis thalami; 5 — пути заднего канатика спинного мозга; 6 — tr. gangliospinothalamicus; 7 — decussatio lemniscorum.
Таким образом, для каждой половины тела в спинном мозге имеются как бы два тракта, передающих импульсы прикосновения: 1) один, неперекрещенный, — в заднем канатике той же стороны и 2) другой, перекрещенный, — в боковом канатике противоположной стороны. Поэтому при одностороннем поражении спинного мозга тактильная чувствительность может оставаться ненарушенной, так как сохраняется соответствующий пучок на здоровой стороне.
В таламусе находится клеточное тело третьего нейрона, аксоны которого направляются в кору большого мозга в составе tr. thalamocorlic?lis, в постцентральную извилину (поля 1, 2, 3) и верхнюю теменную дольку (поля 5, 7), где находится корковый конец кожного анализатора (рис. 350, см. рис. 299).
Тактильная и болевая чувствительность имеет разлитую локализацию в коре головного мозга, что объясняет их меньшее нарушение при ограниченных корковых очагах повреждения.
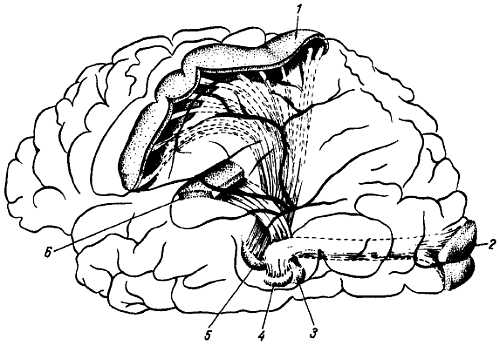
Рис. 350
1 — область общей чувствительности коры полушария; 2 — зрительная область коры полушария; 3 — медиальное коленчатое тело; 4 — латеральное коленчатое тело; 5 — латеральное ядро таламуса; 6 — слуховая область коры головного мозга.
Проводящие пути пространственной кожной чувствительности — стереогноза (узнавание предметов на ощупь) (см. рис. 270). Этот вид кожной чувствительности имеет, как и тактильная чувствительность, идущая по fasc?culus gr?cilis et cune?tus, три звена: 1) спинномозговые ганглии, 2) n?cleus gr?cilis et cune?tus в продолговатом мозге, 3) таламус и, наконец, корковый конец кожного анализатора в верхней теменной дольке (поля 5, 7).
Проводящие пути болевой и температурной чувствительности. Клеточное тело первого нейрона лежит в спинномозговом узле, клетки которого связаны периферическими отростками с кожей, а центральными — с задними рогами спинного мозга (n?clei pr?prii), где помещается клеточное тело второго нейрона (tr?ctus g?ngliospinalis). Аксон второго нейрона переходит на другую сторону в составе commiss?ra ?lba и поднимается в составе tr?ctus spinothal?micus later?lis до таламуса. Следует отметить, что tr?ctus spinothal?micus later?lis в свою очередь делится на две части — переднюю и заднюю, из которых по передней передается болевая чувствительность, а по задней — термическая. В таламусе лежит клеточное тело третьего нейрона, отросток которого в составе tr?ctus thalamocortic?lis направляется в кору большого мозга, где заканчивается в постцентральной извилине (корковый конец кожного анализатора).
Некоторые полагают, что чувство боли воспринимается не только в коре, но и в таламусе, где различные виды чувствительности приобретают эмоциональную окраску. Болевые и температурные импульсы от отделов или органов головы приходят по соответствующим черепным нервам — V, VII, IX, X парам к их чувствительным ядрам и от этих ядер к таламусу и далее в кору постцентральной извилины (нижний отдел).
Вследствие перекреста волокон второго нейрона проводящих путей, идущих от экстероцепторов, импульсы болевой, температурной и частично тактильной чувствительности передаются в постцентральную извилину с противоположной стороны тела. Поэтому следует запомнить, что поражение первого нейрона или второго нейрона до перекреста вызывает расстройство чувствительности на стороне поражения. Если же пострадали волокна второго нейрона после перекреста или третий нейрон, то расстройство тех же видов чувствительности наблюдается на стороне, противоположной очагу поражения.
Проводящие пути от рецепторов внутренних раздражений
Проводящие пути от рецепторов внутренних раздражений могут быть разделены на проводящие пути от аппарата движения (собственно тела), т. е. от проприоцепторов (pr?prius — собственный), составляющих кондуктор двигательного анализатора, и пути от рецепторов внутренностей и сосудов, т. е. интероцепторов; вторая группа путей является кондуктором интероцептивного анализатора.
Проводящие пути двигательного анализатора
Двигательный анализатор воспринимает глубокую проприоцептивную чувствительность, к которой относятся мышечно-суставное чувство, вибрационная чувствительность, чувство давления и веса (гравитация). Основной вид проприоцептивной чувствительности — это мышечно-суставное чувство, т. е. импульсы, которые возникают в связи с изменениями степени натяжения суставной сумки, сухожилий и напряжения мышц; благодаря этим импульсам у человека создается представление о положении тела и частей его в пространстве и об изменении этого положения (что имеет, в частности, значение при полетах в космос, где создается состояние невесомости).
Проводящими путями двигательного анализатора являются tr?ctus gangliobulbothalamocortic?lis и tr?ctus spinocerebell?ris ant?rior et post?rior.
Tr?ctus gangliobulbothalamocortic?lis (см. рис. 349). Рецепторы находятся в костях, мышцах, сухожилиях, суставах, т. е. в собственно теле, отчего называются проприоцепторами (рис. 351).
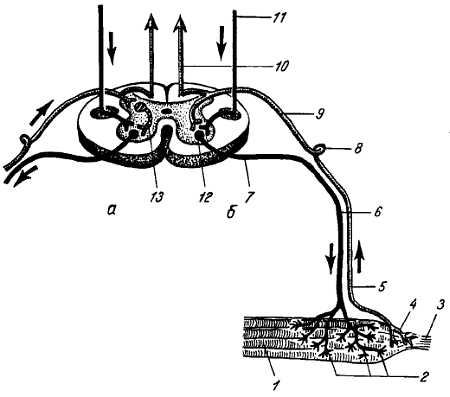
Рис. 351.
1 — мышечные волокна; 2 — двигательные окончания; 3 — сухожилие; 4 — чувствительное окончание в сухожилии; 5 — периферическое волокно чувствительного нейрона; 6 — двигательное волокно; 7 — передний корешок; 8 — спинномозговой узел; 9 — задний корешок; 10 — восходящий чувствительный путь заднего канатика; 11 — нисходящий двигательный путь (боковой пирамидный путь); 12 — двигательная клетка переднего рога; 13 — вставочный нейрон.
Кондуктор состоит из трех нейронов. Клеточное тело первого нейрона помещается в спинномозговом узле. Аксон этой клетки делится на две ветви — периферическую, идущую в составе мышечного нерва от проприоцептора, и центральную, идущую в составе задних корешков в задние канатики спинного мозга, fasciculus gr?cilis и fasciculus cune?tus, до продолговатого мозга (см. рис. 270, 348, 349). Здесь они оканчиваются в соименных ядрах названных канатиков — n?cleus gr?cilis и nucleus cune?tus (tr?ctus gangliobulb?ris).
В этих ядрах помещаются тела вторых нейронов. Аксоны их в составе lemn?scus medi?lis достигают латеральных ядер таламуса, где начинается третье звено. Аксоны клеток последнего направляются через c?psula int?rna (см. рис. 297) в кору предцентральной извилины, где помещается корковый конец двигательного анализатора (поля 4, 6; см. рис. 299). По проприоцептивным путям (пройдя спинномозговые нервы) в кору головного мозга поступают нервные импульсы: по fasc?culus gr?cilis — от мускулатуры нижних конечностей и нижней половины туловища и по fascic?lus cune?tus — от верхней половины туловища и верхней конечности. Проприоцептивные волокна от мышц головы проходят по черепным нервам: тройничному (V) — от мышц глаза и от жевательной мускулатуры, VII — от мимической мускулатуры, IX, X, XI и XII — от языка, от мускулатуры глотки и других мышц бывшего жаберного аппарата.
При выпадении глубокой (проприоцептивной) чувствительности больной утрачивает представление о положении частей своего тела в пространстве и о перемене положения; движения теряют свою четкость, согласованность, наступает расстройство координации движений — атаксия. В отличие от мозжечковой (двигательной) атаксии она называется сенсорной (чувствительной).
Не все пути проприоцептивной чувствительности доходят до коры. Подсознательные проприоцептивные импульсы направляются в мозжечок, который является важнейшим центром проприоцептивной чувствительности.
Проприоцептивные пути к мозжечку (рис. 352). Чувствительные подсознательные импульсы от аппарата движения (костей, суставов, мышц и сухожилий) достигают мозжечка посредством спинальных, проприоцептивных путей, из которых главнейшие — tr?ctus spinocerebell?ris post?rior et ant?rior (см. рис. 270).
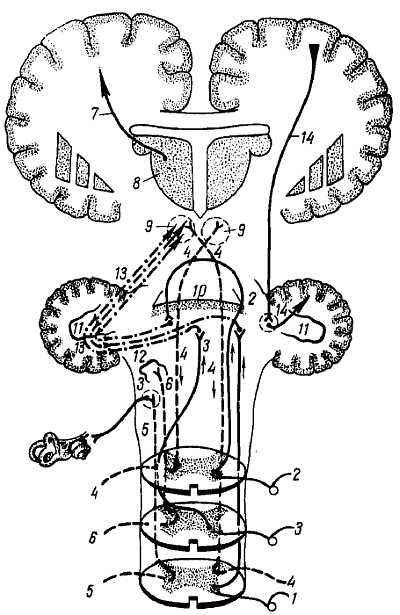
Рис. 352
1 — tr. spinocerebellaris posterior; 2, 3 — tr. spinocerebellaris anterior; 2 — часть, перекрещивающаяся в среднем мозге; 3 — часть, перекрещивающаяся в спинном мозге; 4 — tr. rubrospinalis; 5 — tr. vestibulospinalis; 6 — tr. olivospinalis; 7 — tr. thalamocorticalis; 8 — thalamus; 9 — nucl. ruber; 10 — червячок мозжечка; 11 — nucl. dentatus cerebelli; 12 — олива и ядра pars vestibularis VIII пары черепных нервов; 13 — связи мозжечка и красного ядра; 14 — tr. corticopontocerebellaris.
1. Tr?ctus spinocerebell?ris post?rior. Клеточное тело первого нейрона лежит в спинномозговом узле, аксон делится на две ветви, из которых периферическая идет в составе мышечного нерва от рецептора, заложенного в той или иной части аппарата движения, а центральная в составе заднего корешка проникает в задние рога спинного мозга и при помощи своих концевых ветвей и коллатералей разветвляется вокруг n?cleus thor?cicus. В n?cleus thor?cicus лежат клетки второго нейрона, аксоны которых и образуют tr?ctus spinocerebell?ris post?rior. N?cleus thor?cicus, как показывает название, лучше выражено в грудном отделе на уровне от последнего шейного сегмента до II поясничного. Дойдя в составе бокового канатика спинного мозга до продолговатого по своей стороне, этот тракт в составе нижних мозжечковых ножек достигает коры червя. На своем пути в спинном и продолговатом мозге он не перекрещивается, отчего его называют прямым мозжечковым трактом. Однако, войдя в мозжечок, он большей своей частью перекрещивается в черве.
2. Tr?ctus spinocerebell?ris ant?rior. Первый нейрон тот же, что и у заднего тракта. В subst?ntia interm?dia centr?lis серого вещества спинного мозга помещаются клетки вторых нейронов, аксоны которых, образуя tr?ctus spinocerebell?ris ant?rior, входящий в передние отделы бокового канатика своей и противоположной стороны через commiss?ra ?lba, совершают в ней перекрест. Тракт поднимается через продолговатый мозг и мост до верхнего мозгового паруса, где снова происходит перекрест. После этого волокна входят в мозжечок через его верхние ножки, где заканчиваются в коре червя. В результате весь этот путь оказывается перекрещенным дважды; вследствие этого проприоцептивная чувствительность передается на ту же сторону, с которой поступила.
Таким образом, оба мозжечковых пути соединяют одноименные половины спинного мозга и мозжечка.
Мозжечок получает также проприоцептивные импульсы от n?cleus gr?cilis и n?cleus cune?tus, расположенных в продолговатом мозге. Отростки клеток, заложенных в этих ядрах, идут в мозжечок через его нижние ножки. Все пути глубокой (подсознательной) чувствительности заканчиваются в черве, т. е. в древней части мозжечка, paleocereb?llum.
Интероцептивный анализатор
Интероцептивный анализатор в отличие от других не имеет компактной и морфологически строго очерченной проводниковой части, хотя он и сохраняет специфичность на всем своем протяжении.
Рецепторы его, называемые интероцепторами, рассеяны во всех органах растительной жизни; внутренностях, сосудах, в непроизвольной мускулатуре и железах кожи и др.
Кондуктор состоит из афферентных волокон вегетативной нервной системы, идущих в составе симпатических, парасимпатических и анимальных нервов и далее в спинном и головном мозге до коры. Часть кондуктора интероцептивного анализатора составляют афферентные волокна, идущие в составе черепных нервов (V, VII, IX, X) и несущие импульсы от органов растительной жизни, расположенных в области распространения иннервации каждого из этих нервов. Образуемый ими афферентный путь разбивается на 3 звена: клетки первого звена лежат в узлах этих нервов (g?nglion trigemin?le, g?nglion gen?culi, g?nglion inf?rius); клетки второго нейрона находятся в ядрах этих нервов (n?cleus spin?lis n. trig?mini, n?cleus solit?rius nn. VII, IX, X). Исходящие из этих ядер волокна переходят на другую сторону, направляясь к таламусу. Наконец, клетки третьего звена заложены в таламусе.
Значительную часть кондуктора интероцептивного анализатора образует блуждающий нерв, являющийся главным компонентом парасимпатической иннервации. Идущий по нему афферентный путь также разбивается на 3 звена: клетки первых нейронов лежат в g?nglion inf?rius n. v?gi, клетки вторых нейронов — в n?cleus solit?rius.
Исходящие из этого ядра волокна блуждающего нерва вместе с отростками вторых нейронов языкоглоточного нерва переходят на противоположную сторону, перекрещиваясь с волокнами противоположной стороны, и поднимаются по стволовой части мозга. На уровне верхних холмиков крыши среднего мозга они присоединяются ко вторым нейронам кожного анализатора (lemn?scus medi?lis) и достигают таламуса, где лежат клетки третьих нейронов. Отростки последних идут через заднюю треть задней ножки внутренней капсулы к нижнему отделу постцентральной извилины.
В этом месте располагается одна из частей коркового конца интероцептивного анализатора, связанного с парасимпатическими волокнами черепных нервов и областью их иннервации.
Афферентные пути от органов растительной жизни идут также в составе задних корешков спинномозговых нервов. Клетки первых нейронов в этом случае лежат в спинномозговых узлах. Мощный коллектор афферентного пути от органов растительной жизни проходит через внутренностные нервы (nn. spl?nchnici m?jor et m?nor). Различные группы нервных волокон этих нервов восходят в спинном мозге в составе его задних и боковых канатиков. Афферентные волокна задних канатиков передают интероцептивные импульсы, достигающие через таламусы коры большого мозга.
Афферентные волокна боковых канатиков оканчиваются в ядрах ствола мозга, мозжечка и таламуса (n?cleus ventr?lis post?rior). Итак, в таламусе лежат клетки третьих нейронов всего кондуктора интероцептивного анализатора, связанного как с симпатической, так и с парасимпатической иннервацией. Поэтому в таламусе происходит замыкание интероцептивных рефлекторных дуг и возможен «выход» на эфферентные пути.
Замыкание для отдельных рефлексов может происходить и на других, более низких уровнях. Этим объясняется автоматическая, подсознательная, деятельность органов, управляемых вегетативной нервной системой. Корковый конец интероцептивного анализатора, кроме постцентральной извилины, находится в премоторной зоне, где заканчиваются афферентные волокна, идущие от таламуса. Интероцептивные импульсы идут по внутренностным нервам, достигают также коры пред- и постцентральных извилин в зонах кожно-мышечной чувствительности.
Возможно, что эти зоны содержат первые корковые нейроны эфферентных путей вегетативной нервной системы, осуществляющие кортикальную регуляцию вегетативных функций. С этой точки зрения эти первые корковые нейроны могут рассматриваться как своего рода аналоги пирамидных клеток, являющихся первыми нейронами пирамидных путей.
Как видно из вышеизложенного, интероцептивный анализатор в структурном и функциональном отношениях сходен с экстероцептивными анализаторами, однако площадь коркового конца интероцептивного анализатора значительно меньше по сравнению с экстероцептивными. Этим объясняется его «грубость», т. е. меньшая тонкость, точность дифференцировок по отношению к сознанию.
На всех уровнях центральной нервной системы: в спинном мозге, мозжечке, в таламусах и коре большого мозга — имеется весьма тесное перекрытие путей и зон представительства анимальных и вегетативных органов. Висцеральные и соматические афферентные импульсы могут адресоваться к одному и тому же нейрону, «обслуживающему» и вегетативные, и соматические функции. Все это обеспечивает взаимодействие анимальной и вегетативной частей единой нервной системы. Высшая интеграция анимальных и вегетативных функций осуществляется в коре головного мозга, особенно в премоторной зоне.
До сих пор были рассмотрены афферентные пути, связанные с определенной специализацией нейронов, проводящие те или иные специфические импульсы (тактильные, проприоцептивные, интероцептивные). Вместе с проводящими путями от органов зрения, слуха, вкуса, обоняния они составляют так называемую специфическую афферентную систему. Наряду с этим существует афферентная система, представленная так называемой ретикулярной формацией, относящаяся к неспецифическим структурам. Ретикулярная формация воспринимает все без исключения импульсы: болевые, световые, звуковые и т. д. Но в то время как специфические импульсы от каждого органа чувств поступают по специальным проводниковым системам в кору соответствующих анализаторов, в ретикулярной формации не существует специализации нейронов; одни и те же нейроны воспринимают различные импульсы и передают их во все слои коры. Таким образом, ретикулярная формация составляет вторую афферентную систему.