ВТОРАЯ АФФЕРЕНТНАЯ СИСТЕМА ГОЛОВНОГО МОЗГА — РЕТИКУЛЯРНАЯ ФОРМАЦИЯ, FORMATIO RETICULARIS
Под этим названием подразумевают совокупность структур, расположенных в центральных отделах мозгового ствола и отличающихся следующими морфологическими особенностями:
1. Нейроны ретикулярной формации имеют отличающее их от других нейронов строение: дендриты их ветвятся очень слабо, нейриты, наоборот, делятся на восходящую и нисходящую ветви, которые отпускают от себя многочисленные коллатерали, благодаря чему аксон может контактировать с огромным числом нервных клеток (при длине в 2 см — с 27 500).
2. Нервные волокна идут в самых различных направлениях, напоминая под микроскопом сеть, что и послужило основанием для Дейтерса назвать ее 120 лет тому назад сетчатой, или ретикулярной, формацией.
3. Клетки ретикулярной формации местами рассеяны, а местами образуют ядра, начало выделению которых положил В. М. Бехтерев, описавший ретикулярное ядро покрышки моста (n?cleus reticul?ris tegm?nti p?ntis).
В настоящее время описано 96 отдельных ядер.
Область распространения ретикулярной формации точно еще не установлена. На основании физиологических данных, она расположена по всей длине мозгового ствола и занимает центральное положение в продолговатом мозге, мосте, среднем мозге, в гипоталамической области и даже в медиальной части таламусов. Как филогенетически более древняя ретикулярная формация локализуется в покрышке ствола головного мозга.
Связи ретикулярной формации. Ретикулярная формация связана со всеми отделами центральной нервной системы. Различают: 1) ретикулопетальные связи, идущие от всех отделов головного мозга; 2) ретикулофугальные связи, идущие к серому веществу и ядрам головного и спинного мозга; 3) ретикулоретикулярные связи (восходящие и нисходящие) между различными ядрами самой ретикулярной формации.
Функция. В настоящее время считают, что ретикулярная формация является «генератором энергии» и регулирует процессы, совершающиеся в других отделах центральной нервной системы, включая и кору большого мозга. Особенно важно, что ретикулярная формация оказывает общее (генерализованное) неспецифическое активизирующее воздействие на всю кору головного мозга, (П. К. Анохин), что обеспечивается наличием восходящих про водящих путей от сетчатой формации ко всем долям мозговых полушарий. Поэтому ее называют также восходящей активирующей ретикулярной системой. Будучи связана коллатералями аксонов своих клеток со всеми проходящими через ствол мозга специфическими афферентными проводящими путями, она получает от них импульсы и несет неспецифическую информацию в мозговую кору.
В результате через мозговой ствол проходят в кору мозга две афферентные системы: одна специфическая — это все специфические чувствительные проводящие пути, несущие импульсы от всех рецепторов (экстероинтеро- и проприоцепторов) и заканчивающиеся на телах клеток преимущественно IV слоя коры; другая — неспецифическая, образованная ретикулярной формацией и заканчивающаяся на дендритах всех слоев коры.
Взаимодействие обеих этих систем обусловливает окончательную реакцию корковых нейронов. Таково современное представление о двух афферентных системах головного мозга.
Учитывая столь большое значение ретикулярной формации и ее влияние на кору мозга, некоторые зарубежные исследователи преувеличивают ее роль, считая, что она, располагаясь в центральных частях мозга, составляет особую «центрэнцефалическую» систему, выполняющую функцию сознания и интеграции. Стремление спустить высший уровень интеграции из коры мозга в подкорку не имеет под собой фактических оснований и является антиэволюционным, так как в процессе эволюции наибольшего развития достигает наивысший отдел мозга, т. е. его плащ, а не ствол. Это стремление противоречит материалистической идее нервизма и отражает фрейдизм — идеалистическое учение о ведущей роли не коры, а подкорки. Строение и функция ретикулярной формации полностью еще не раскрыты и составляют предмет дальнейших изысканий.
ЭФФЕРЕНТНЫЕ (НИСХОДЯЩИЕ) ПРОВОДЯЩИЕ ПУТИ
Нисходящие двигательные пути идут от коры головного мозга — tr?ctus corticonucle?ris et corticospin?lis (пирамидная система), от подкорковых ядер переднего мозга — экстрапирамидная система и от мозжечка.
Корково-спинномозговой (пирамидный) путь, или пирамидная система.
Клеточное тело первого нейрона лежит в предцентральной извилине коры большого мозга (гигантские пирамидные клетки). Аксоны этих клеток через cor?na radi?ta спускаются во внутреннюю капсулу (колено и передние две трети задней ножки), далее в b?sis ped?nculi c?rebri (срединный ее отдел), а затем в pars basil?ris моста и продолговатый мозг. Здесь часть волокон пирамидной системы вступает в связь с ядрами черепных нервов. Эта часть пирамидной системы, проходящая через колено внутренней капсулы и связывающая кору большого мозга с ядрами черепных нервов, называется tr?ctus corticonucle?ris[38]. Волокна этого тракта частью переходят на другую сторону, частью остаются на своей стороне. Аксоны клеток, заложенных в ядрах черепных нервов (клеточные тела вторых нейронов), в составе соответственных нервов оканчиваются в скелетной мускулатуре, иннервируемой этими нервами.
Другая часть пирамидной системы, проходящая в передних двух третях задней ножки внутренней капсулы, служит для связи с ядрами спинно-мозговых нервов, спускается до передних рогов спинного мозга и потому называется tr?ctus corticospin?lis. Этот тракт, пройдя в мозговом стволе до продолговатого мозга, образует в нем пирамиды. В последних перекрещивается часть волокон tr?ctus corticospin?lis (decuss?tio pyram?dum), которая, спускаясь в спинной мозг, ложится в боковой его канатик, образуя, tr?ctus corticospin?lis (pyramid?lis) later?lis. Оставшаяся неперекрещенной часть tr?ctus corticospin?lis спускается в переднем канатике спинного мозга, образуя его tr?ctis corticospin?lis (pyramid?lis) ant?rior (см. рис. 270).
Волокна этого пучка постепенно по протяжению спинного мозга также переходят на другую сторону в составе commiss?ra ?lba, в результате чего весь tr?ctus corticospin?lis оказывается перекрещенным. Благодаря этому кора каждого полушария иннервирует мускулатуру противоположной стороны тела.
Двигательные и чувствительные перекресты, происходящие в различных отделах мозга (decuss?tio pyram?dum, commiss?ra ?lba, decuss?tio lemnisc?rum и др.), представляют, по И. П. Павлову, приспособление нервной системы, направленное на сохранение иннервации при повреждении мозга в каком-либо месте одной его стороны. Аксоны, составляющие tr?ctus corticospin?lis (pyramid?lis), вступают в связь с двигательными клетками передних рогов спинного мозга, где начинается второе звено[39]. Аксоны лежащих здесь клеток идут в составе передних корешков и далее мышечных нервов к скелетной мускулатуре туловища и конечностей, иннервируемой спинно-мозговыми нервами. Таким образом, tr?ctus corticonucle?ris и tr?ctus corticospin?lis составляют единую пирамидную систему, служащую для сознательного управления скелетной мускулатурой (рис. 353). Эта система особенно развита у человека в связи с прямохождением и сознательным пользованием своим аппаратом движения в процессах труда и членораздельной речью.
Нисходящие пути подкорковых ядер переднего мозга — экстрапирамидная система
Пирамидная система, как уже отмечалось выше, начинается в коре большого мозга (V слой, пирамидные клетки). Экстрапирамидная система (рис. 354, 355) слагается из подкорковых образований. В ее состав входят c?rpus stri?tum, thal?mus, n?cleus hypothal?micus post?rior, n?cleus r?ber, subst?ntia n?gra и связывающие их проводники белого вещества. Экстрапирамидная система отличается от пирамидной по своему развитию, строению и функции. Она является старейшим в филогенетическом отношении моторно-тоническим аппаратом, который встречается уже у рыб, у которых имеется еще только бледный шар, pall?dum (paleostri?tum), у амфибий появляется уже скорлупа, put?men (neostri?tum).
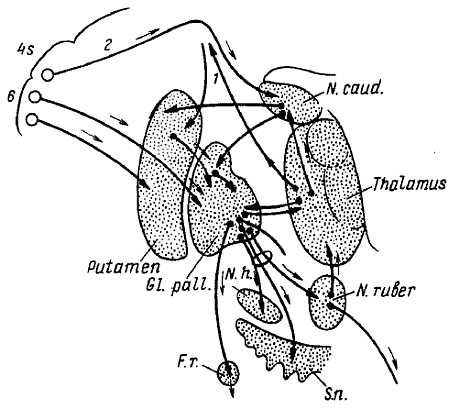
Рис. 354.
6—4s — поля премоторной и двигательной зоны коры мозга; 1 — волокна, восходящие из таламуса в кору; 2 — путь от «тормозных» участков поля 4s в хвостатое ядро (N. caud); Gl. pall. — бледный шар; N. h. — гипоталамическое ядро; N. ruber — красное ядро; S. n. — черная субстанция; F. r. — ретикулярная формация продолговатого мозга. Стрелки указывают направление и «станцию назначения» импульсов.
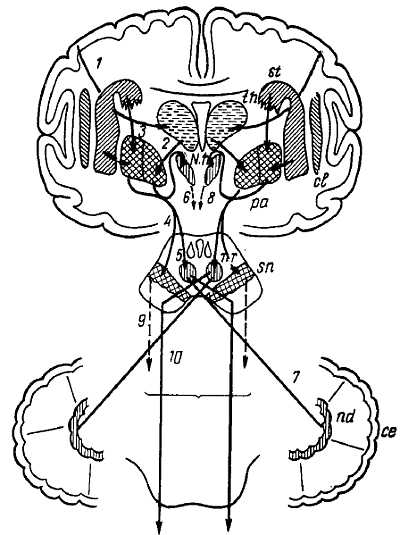
Рис. 355
се — кора мозжечка; сl — claustrum; N. h. — гипоталамическое ядро; nd — nucl. dentatus cerebelli; nr — nucl. ruber; pa — pallidum; sn — substantia nigra; st — striatum (nucl. caudatus и putamen); th — thalamus; 1 — tr. corticostrialis; 2 — fibrae thalamopallidales: 3 — fibrae strioppallidales; 4, 5 — связи с substantia nigra и nucl. ruber; 6, 8 — эфферентные волокна гипоталамического ядра; 7 — волокна pedunculus cerebellaris superior; 9 — эфферентные волокна substantia nigra; 10 — tr. rubrospinalis.
На этой стадии развития, когда пирамидная система еще отсутствует, экстрапирамидная система является высшим отделом головного мозга, воспринимающим раздражение от рецепторов органов и посылающим импульсы к мускулатуре через автоматические механизмы спинного мозга. В результате возникают сравнительно простые движения (автоматизированные). У млекопитающих по мере развития переднего мозга и его коры образуется новая кинетическая система — пирамидная, соответствующая новой форме двигательных актов, связанных со все большей специализацией небольших групп мышц. В результате у человека в полной мере развиваются две системы:
1. Пирамидная система — филогенетически более молодая, представлена экранными центрами коры, ведающими сознательными движениями человека. Через пирамидную систему осуществляется также в движениях корковая деятельность, основанная на условных рефлексах.
2. Экстрапирамидная система — филогенетически более старая, состоящая из подкорковых ядер. У человека она играет подчиненную роль и осуществляет высшие безусловные рефлексы, поддерживая тонус мускулатуры и автоматически регулируя ее работу (непроизвольная автоматическая иннервация скелетной мускулатуры). Эта автоматическая регуляция мышц осуществляется благодаря связям всех компонентов экстрапирамидной системы между собой и с n?cleus r?ber, от которого идет нисходящий двигательный путь к передним рогам серого вещества спинного мозга, tr?ctus rubrospin?lis. Этот тракт начинается в клетках красного ядра, переходит через срединную плоскость на уровне верхних холмиков крыши среднего мозга, образуя вентральный перекрест (decuss?tio ventr?lis tegm?nti), и спускается через мозговой ствол в боковые канатики спинного мозга, после чего заканчивается на двигательных нейронах передних рогов серого вещества. Таким образом, экстрапирамидная система действует на спинной мозг через красное ядро, которое составляет важнейшую часть этой системы.
К работе экстрапирамидной системы имеют отношение нисходящие мозжечковые пути, а также ретикулярно-спинномозговой путь, которому в настоящее время придается большое значение в регуляции двигательной активности спинного мозга.
Нисходящие двигательные пути мозжечка
Мозжечок принимает участие в контроле двигательных нейронов спинного мозга (мышечная координация, поддержание равновесия, сохранение мышечного тонуса и преодоление инерции и силы тяжести). Это осуществляется с помощью tr?ctus cerebellorubrospin?lis (см. рис. 352). Клеточное тело первого звена этого пути лежит в коре мозжечка (грушевидные нейроциты). Их аксоны заканчиваются в n?cleus dent?tus cereb?lli и, возможно, в других ядрах мозжечка, где начинается второе звено. Аксоны вторых нейронов идут через верхние мозжечковые ножки к среднему мозгу и оканчиваются в n?cleus r?Ьег. Здесь помещаются клетки третьего звена, аксоны которых в составе tr?ctus rubrospin?lis, переключившись в двигательных нейронах передних рогов спинного мозга (четвертое звено), достигают скелетной мускулатуры.
Нисходящие пути коры большого мозга к мозжечку
Кора большого мозга, ведающая всеми процессами организма, держит в своем подчинении и мозжечок как важнейший проприоцептивный центр, связанный с движениями тела. Это достигается наличием специального нисходящего пути от коры большого мозга к коре мозжечка — tr?ctus corticopontocerebell?ris (см. рис. 352).
Первое звено этого пути состоит из нейронов, клеточные тела которых заложены в коре большого мозга, а аксоны спускаются к ядрам моста, n?clei (pr?prii) p?ntis. Эти нейроны составляют отдельные пучки, которые соответственно различным долям мозга называются tr?ctus frorttopont?nus, occipitopont?nus, temporopont?nus et parietopont?nus. В ядрах моста начинаются вторые нейроны, аксоны которых образуют tr?ctus роntocerebell?ris, идущий на противоположную сторону моста, и в составе средних мозжечковых ножек он достигает коры полушарий мозжечка (neocerebellum).
Таким образом, устанавливается связь между корой большого мозга и полушариями мозжечка. (Полушария головного мозга связаны с противоположными полушариями мозжечка.) Оба эти отдела головного мозга являются более молодыми и в своем развитии взаимосвязаны. Чем сильнее развиты кора и полушария большого мозга, тем сильнее развиты кора и полушария мозжечка. Так как связь этих отделов головного мозга осуществляется через мост, то и степень развития последнего определяется развитием мозговой коры.
Следовательно, три пары ножек мозжечка обеспечивают его многосторонние связи: через нижние ножки он получает импульсы из спинного мозга и продолговатого мозга, через средние — из коры полушарий большого мозга; в составе верхних ножек проходит главный эфферентный путь мозжечка, по которому мозжечковые импульсы передаются на клетки передних рогов спинного мозга. Связь полушарий головного мозга с полушариями мозжечка, т. е. с его новой частью (neocereb?llum), перекрестная, связь же червя, т. е. старой части мозжечка (paleocereb?llum), со спинным мозгом главным образом прямая, гомолатеральная.


![Дочки-матери [Все, о чем вам не рассказывала ваша мама и чему стоит научить свою дочь]](https://img-lib.med-tutorial.ru/3386939452/cover.jpg)



