Периферический отдел нервной системы
АНИМАЛЬНЫЕ ИЛИ СОМАТИЧЕСКИЕ НЕРВЫ
По месту отхождения от центральной нервной системы — от спинного или головного мозга, нервные стволы разделяются на спинномозговые, nn. spin?les, и черепные, nn. crani?les (enceph?lici).
СПИННОМОЗГОВЫЕ НЕРВЫ
Спинномозговые нервы, nn. spin?les, располагаются в правильном порядке (невромеры), соответствуя миотомам (миомерам) туловища и чередуясь с сегментами позвоночного столба; каждому нерву соответствует относящийся к нему участок кожи (дерматом).
У человека имеется 31 пара спинномозговых нервов, а именно: 8 пар шейных, 12 пар грудных, 5 пар поясничных, 5 пар крестцовых и 1 пара копчиковых (см. рис. 267). Каждый спинномозговой нерв отходит от спинного мозга двумя корешками: задним (чувствительным) и передним (двигательным); оба корешка соединяются в один ствол, tr?ncus n. spin?lis, выходящий из позвоночного канала через межпозвоночное отверстие. Вблизи и несколько кнаружи от места соединения задний корешок образует узел, g?nglion spin?le, в котором передний двигательный корешок не принимает участия. Благодаря соединению обоих корешков спинномозговые нервы являются смешанными нервами: они содержат чувствительные (афферентные) волокна от клеток спинномозговых узлов, двигательные (эфферентные) волокна от клеток переднего рога, а также вегетативные волокна от клеток боковых рогов, выходящие из спинного мозга в составе переднего корешка (рис. 308).
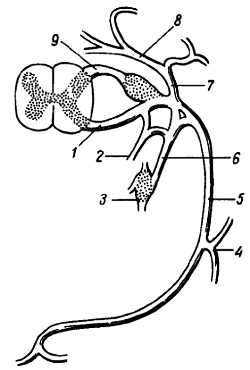
Рис. 308
1 — передний корешок: 2 — r. meningeus; 3 — симпатический узел; 4 — r. cutaneus lateralis; 5 — r. ventralis; 6 — r. communicans; 7 — r. dorsalis; 8 — r. medialis rami dorsalis; 9 — задний корешок.
Вегетативные волокна имеются и в заднем корешке. Вегетативные волокна, попадающие через корешки в анимальные нервы, обеспечивают в соме такие процессы, как трофика, сосудодвигательные реакции и т. п.
У круглоротых (миноги) оба корешка продолжаются в отдельные нервы — двигательные и чувствительные. В дальнейшем ходе эволюции, начиная с поперечноротых рыб корешки сближаются и сливаются, так что раздельный ход сохраняется только для корешков, а нервы становятся смешанными.
Каждый спинномозговой нерв при выходе из межпозвоночного отверстия делится соответственно двум частям миотома (дорсальной и вентральной) на две ветви:
1) заднюю, r?mus dors?lis, для развивающейся из дорсальной части миотома аутохтонной мускулатуры спины и покрывающей ее кожи;
2) переднюю, r?mus ventr?lis, для вентральной стенки туловища и конечностей, развивающихся из вентральных частей миотомов.
Кроме того, от спинномозгового нерва отходят еще два рода ветвей:
3) для иннервации внутренностей и сосудов — соединительные ветви к симпатическому стволу, rr. coinmunic?ntes;
4) для иннервации оболочек спинного мозга — r. men?ngeus, идущая обратно через межпозвоночное отверстие.
Задние ветви спинномозговых нервов
Задние ветви, r?mi dors?les, всех спинномозговых нервов идут назад между поперечными отростками позвонков, огибая суставные отростки их. Все они (за исключением I шейного, IV и V крестцовых и копчикового) делятся на r?mus medi?lis и r?mus later?lis, которые снабжают кожу затылка, задней поверхности шеи и спины, а также глубокие спинные мышцы.
Задняя ветвь I шейного нерва, n. suboccipit?lis, выходит между затылочной костью и атлантом и затем делится на ветви, снабжающие mm. r?cti c?pitis m?jor et m?nor, m. semispin?lis c?pitis, mm. obl?qui c?pitis. К коже n. suboccipit?lis ветвей не дает. Задняя ветвь II шейного нерва, n. occipit?lis m?jor, выйдя между задней дугой атланта и II позвонком, прободает затем мышцы и, сделавшись подкожным, иннервирует затылочную область головы.
R?mi dors?les грудных нервов делятся на медиальную и латеральную ветви, дающие ветви к аутохтонной мускулатуре; кожные ветви у верхних грудных нервов отходят только от r?mi medi?дes, а у нижних — от r?mi later?les.
Кожные ветви трех верхних поясничных нервов идут в верхнюю часть ягодичной области под названием nn. cl?nium superi?res, а кожные ветви крестцовых — под названием nn. cl?nium m?dii.
Передние ветви спинномозговых нервов
Передние ветви, r?mi ventr?les, спинномозговых нервов иннервируют кожу и мускулатуру вентральной стенки тела и обе пары конечностей. Так как кожа живота в нижней своей части принимает участие в развитии наружных половых органов, то покрывающая их кожа иннервируется также передними ветвями. Последние, кроме первых двух, гораздо крупнее задних.
Передние ветви спинномозговых нервов сохраняют первоначальное метамерное строение только в грудном отделе (nn. intercost?les). В остальных отделах, связанных с конечностями, при развитии которых сегментарность теряется, волокна, отходящие от передних спинномозговых ветвей, переплетаются. Так образуются нервные сплетения, pl?xus, в которых происходит обмен волокон различных невромеров. В сплетениях происходит сложное перераспределение волокон: передняя ветвь каждого спинномозгового нерва дает свои волокна в несколько периферических нервов, и, следовательно, каждый из них содержит волокна от нескольких сегментов спинного мозга. Понятно поэтому, что поражение того или иного нерва, не сопровождается нарушением функции всех мышц, получающих иннервацию из сегментов, давших начало этому нерву.
Большинство нервов, отходящих от сплетений, являются смешанными; поэтому клиническая картина поражения складывается из двигательных нарушений, нарушений чувствительности и вегетативных расстройств.
Различают три больших сплетения: шейное, плечевое и пояснично-крестцовое. Последнее делится на поясничное, крестцовое и копчиковое.
Шейное сплетение
Шейное сплетение, pl?xus cervic?lis, образуется передними ветвями четырех верхних шейных нервов (СI — CIV), которые соединяются между собой тремя дугообразными петлями и располагаются сбоку поперечных отростков между предпозвоночными мышцами с медиальной и позвоночными (m. scal?nus m?dius, m. lev?tor sc?pulae, m. spl?nius c?rvicis) с латеральной стороны, анастомозируя с n. access?rius, n. hypogl?ssus и truncus sympathicus. Спереди сплетение прикрыто m. sternocleidomastoideus. Ветви, отходящие от сплетения, разделяются на кожные, мышечные и смешанные (рис. 309).
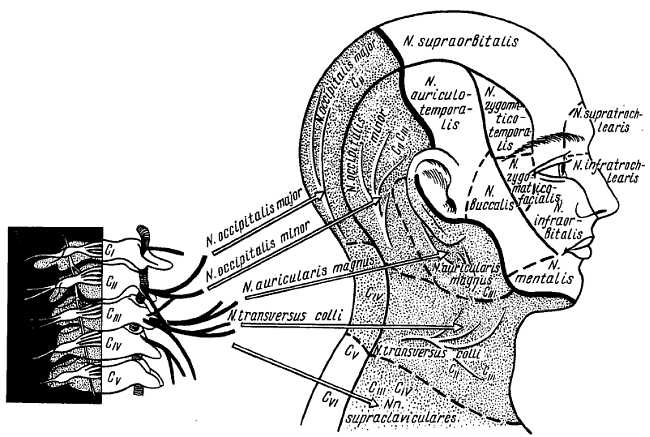
Рис. 309.
Кожные ветви. 1. N. occipit?lis m?nor (из СII и СIII) к коже латеральной части затылочной области.
2. N. auricul?ris m?gnus (из CIII) иннервирует ушную раковину и наружный слуховой проход.
3. N. transv?rsus c?lli (из СII-СIII) отводит, как и предыдущие два нерва у середины заднего края m. sternocle?domastoideus и, обогнув задний край грудино-ключично-сосцевидной мышцы, идет кпереди и снабжает кожу шеи.
4. Nn. supraclavicul?res (из СIII и CIV) спускаются в кожу над большой грудной и дельтовидной мышцами.
Мышечные ветви. 1. К mm. r?cti c?pitis ant?rior et later?lis, mm. l?ngi c?pitis et c?lli, mm. scal?ni, m. lev?tor sc?pulae и, наконец, к mm. intertransvers?rii anteri?res.
2. R?dix inf?rior ?nsae cervic?lis, отходит от СII-СIII, проходит спереди от v. jugul?ris interna под грудино-ключично-сосцевидной мышцей и соединяется с radix superior, отходящим от n. hypogl?ssus, образуя вместе с этой ветвью шейную петлю, ?nsa cervic?lis. Волокна шейного сплетения посредством ветвей, отходящих от ?nsa, иннервируют m. stemohyo?deus, m. stemothyro?deus и m. omohyo?deus.
3. Ветви к m. sternocleidomasto?deus и m. trapezius (от СIII и CIV), принимающие участие в иннервации этих мышц вместе с n. access?rius.
Смешанные ветви. N. phr?nicus — диафрагмальный нерв (СIII-CIV), спускается по m. scal?nus ant?rior вниз в грудную полость, куда проходит между подключичной артерией и веной. Далее правый n. phr?nicus спускается почти вертикально впереди корня правого легкого и идет по боковой поверхности перикарда, к диафрагме. Левый n. phr?nicus пересекает переднюю поверхность дуги аорты и впереди корня левого легкого проходит по левой боковой поверхности перикарда к диафрагме. Оба нерва идут в переднем средостении между перикардом и плеврой. N. phr?nicus принимает волокна от двух нижних шейных узлов симпатического ствола.
N. phr?nicus — смешанный нерв: своими двигательными ветвями он иннервирует диафрагму, являясь, таким образом, нервом, обслуживающим дыхание; чувствительные ветви он дает к плевре и перикарду. Некоторые из конечных ветвей нерва проходят сквозь диафрагму в брюшную полость (nn. phrenicoabdomin?les) и анастомозируют с симпатическим сплетением диафрагмы, посылая веточки к брюшине, связкам печени и к самой печени, вследствие чего при ее заболевании может возникнуть особый френикус-симптом. Своими волокнами в грудной полости он снабжает сердце, легкие, вилочковую железу, а в брюшной он связан с чревным сплетением и через него иннервирует ряд внутренностей.
Плечевое сплетение
Плечевое сплетение, pl?xus brachi?lis, слагается из передних ветвей четырех нижних шейных нервов (СV—СVIII) и большей части первого грудного (ThI); часто присоединяется тонкая ветвь от CIV. Плечевое сплетение выходит через промежуток между передней и средней лестничными мышцами в надключичную ямку, располагаясь выше и сзади a. subcl?via. Из него возникают три толстых нервных пучка, идущих в подмышечную ямку и окружающих a. axill?ris с трех сторон: с латеральной (латеральный пучок), медиальной (медиальный пучок) и кзади от артерии (задний пучок).
В сплетении обыкновенно различают надключичную (pars supraclavicul?ris) и подключичную (pars infraclavicul?ris) части. Периферические ветви разделяются на короткие и длинные. Короткие ветви отходят в различных местах сплетения в надключичной его части и снабжают отчасти мышцы шеи, а также мышцы пояса верхней конечности (за исключением m. trap?zius) и плечевого сустава. Длинные ветви происходят из вышеуказанных трех пучков и идут вдоль верхней конечности, иннервируя ее мышцы и кожу.
Короткие ветви. 1. N. dors?lis sc?pulae (из CV) идет вдоль медиального края лопатки. Иннервирует m. lev?tor sc?pulae и mm. rhombo?dei.
2. N. thor?cicus l?ngus (из CV—CVII) спускается по наружной поверхности m. serr?tus ant?rior, которую иннервирует.
3. N. suprascapul?ris (из CV и CVI) идет через incis?ra sc?pulae в f?ssa supraspin?ta. Иннервирует mm. supra- et infraspin?tus и капсулу плечевого сустава.
4. Nn. pector?les medi?lis et later?lis (из CV—ThI) — к m. pector?lis m?jor et m?nor.
5. N. subcl?vius (CV) — к m. subcl?vius.
6. N. subscapul?ris (CV—СVIII) иннервирует m. subscapui?ris, m. t?res m?jor и m. lat?ssimus d?rsi. Ветвь, идущая вдоль латерального края лопатки к m. lat?ssimus d?rsi, называется n. thoracodors?lis.
7. N. axill?ris, подмышечный нерв (из CV—CVI), — самый толстый нерв из коротких ветвей плечевого сплетения, проникает вместе с a. circumfl?xa h?meri post?rior через for?men quadril?terum на заднюю поверхность хирургической шейки плечевой кости и дает ветви к mm. delto?deus, t?res m?nor и к плечевому суставу. По заднему краю дельтовидной мышцы дает кожную ветвь, n. cut?neus br?chii later?lis sup?rior, иннервирующую кожу дельтовидной области и заднелатеральной области плеча в верхнем отделе его.
Длинные ветви. Среди них можно выделить передние — для сгибателей и пронаторов (nn. musculocutaneus, medi?nus et uln?ris) и задние — для разгибателей и супинаторов (n. radi?lis).
1. N. musculocut?neus, мышечно-кожный нерв, отходит от латерального пучка плечевого сплетения (из CV—CVII), прободает m. coracobrachi?lis и иннервирует все передние мышцы плеча mm. coracobrachi?lis, biceps et brachi?lis. Пройдя между двумя последними на латеральную сторону плеча, продолжается на предплечье под названием n. cut?neus antebr?chii later?lis, снабжая кожу лучевой стороны последнего, а также кожу th?nar.
2. N. medi?nus, срединный нерв (CV—CVIII, ThI), отходит от медиального и латерального пучков двумя корешками, охватывающими спереди a. axill?ris, затем он идет в s?lcus bicipit?lis medi?lis вместе с плечевой артерией. В локтевом сгибе нерв подходит под m. pron?tor t?res и поверхностный сгибатель пальцев и идет дальше между последним и m. fl?xor digit?rum prof?ndus, затем — в одноименной бороздке, s?lcus medi?nus, посередине предплечья на ладонь. На плече n. medi?nus ветвей не дает. На предплечье он отдает r?mi muscul?res для всех мышц передней сгибательной группы, за исключением m. fl?xor c?rpi uln?ris и ближайшей к последнему части глубокого сгибателя пальцев (рис. 310).
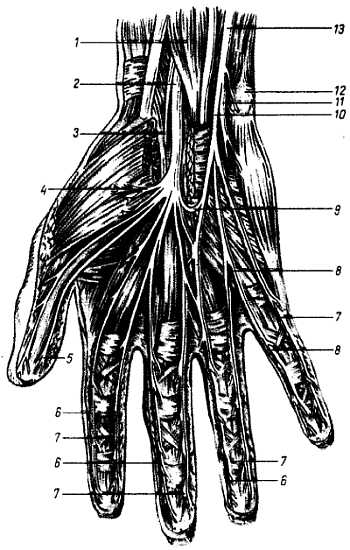
Рис. 310.
1 — m. flexor digitorum superfieialis; 2 — n. medianus; 3 — r. palmaris n. mediani; 4 — rr. musculares n. mediani; 5, 6, 7, 8 — nn. digitales palmares proprii; 9 — r. communicans между n. medianus и n. ulnaris; 10 — r. superfieialis n. ulnaris; 11 — r. profundus n. ulnaris; 12 — os pisiforme; 13 — n. ulnaris.
Одна из ветвей, n. inter?sseus (antebr?chii) ant?rior, сопровождает a. inter?ssea ant?rior на межкостной перепонке, и иннервирует глубокие сгибающие мышцы (m. fl?xor p?llicis l?ngus и часть m. fl?xor digit?rum prof?ndus), m. pron?tor quadr?tus и лучезапястный сустав. Над лучезапястным суставом n. edi?nus дает тонкую кожную ветвь — r?mus palm?ris n. medi?ni, которая снабжает небольшой участок кожи на th?nar и ладони. N. medi?nus выходит на ладонь через can?lis c?rpi вместе с сухожилиями сгибателей и делится на три ветви, nn. digit?les palm?res comm?nes, которые идут вдоль первого, второго и третьего межпястных промежутков под ладонным апоневрозом по направлению к пальцам. Первая из них иннервирует мышцы th?nar, за исключением m. add?ctor p?llicis и глубокой головки m. fl?xor p?llicis br?vis, которые иннервируются локтевым нервом. Nn. digit?les palm?res communes в свою очередь делятся на семь nn. digit?les palm?res pr?prii, которые идут к обоим сторонам I–III пальцев и к лучевой стороне IV пальца. От этих же ветвей снабжается и кожа лучевой стороны ладони (рис. 311); пальцевые нервы снабжают также первую и вторую червеобразные мышцы.
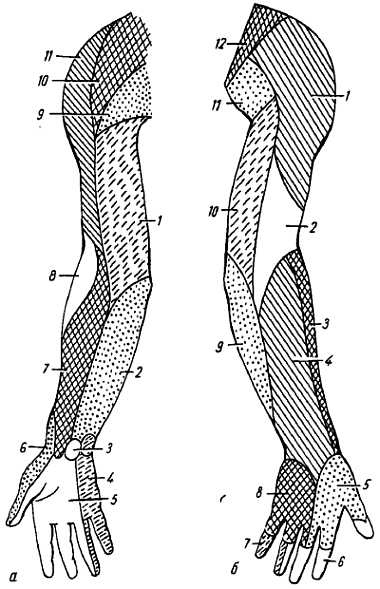
Рис. 311
а — ладонная поверхность: 1 — n. cutaneus brachii medialis; 2 — n. cutaneus antebrachii medialis; 3 — r. palmaris n. mediani; 4 — r. cutaneus palmaris n. ulnaris; 5 — n. medianus; 6 — r. superificialis n. radialis; 7 — n. cutaneus antebrachii lateralis (от n. musculocutaneus); 8 — n. cutaneus brachii posterior (от n. radialis); 9 — nn. pectorales laterales; 10 — nn. supraclaviculares (от plexus cervicalis); 11 — n. cutaneus brachii lateralis superior (от n. axillaris);
б — тыльная поверхность: 1 — n. cutaneus brachii lateralis superior (от n. axillaris); 2 — n. cutaneus brachii posterior et lateralis inferior (от n. radialis); 3 — n. cutaneus antebrachii lateralis (от n. musculocutaneus); 4 — n. cutaneus antebrachii posterior (от n. radialis); 5 — r. superficial n. radialis; 6 — n. medianus; 7 — r. cutaneus palmaris n. ulnaris; 8 — r. dorsalis n. ulnaris; 9 — n. cutaneus antebrachii medialis; 10 — n. cutaneus brachii medialis; 11 — rr. cutanet latt. nn. intercostalium; 12 — nn. supraclaviculares (от plexus cervicalis).
3. N. uln?ris, локтевой нерв (см. рис. 310, 311), выходящий из медиального пучка плечевого сплетения (СVII, СVIII, ThI), проходит по медиальной стороне плеча на заднюю поверхность медиального надмыщелка (здесь он лежит под кожей, почему его часто ушибают, что вызывает в медиальной зоне предплечья ощущение покалывания), затем ложится в s?lcus uln?ris и далее в can?lis c?rpi uln?ris, где идет вместе с соименными артерией и венами до ладони; на поверхности retin?culum flex?rum он переходит в свою конечную ветвь — r?mus palm?ris n. uln?ris. На плече локтевой нерв, так же как и срединный, не дает ветвей.
Ветви n. uln?ris на предплечье и кисти. R?mi articul?res к локтевому суставу.
R?mi muscul?res для m. fl?xor c?rpi uln?ris и соседней с ним части m. fl?xor digit?rum prof?ndus.
R?mus cut?neus palm?ris к коже hypoth?nar.
R?mus dors?lis n. uln?ris уходит через промежуток между m. fl?xor c?rpi uln?ris и локтевой костью на тыл кисти, где она разделяется на пять тыльных пальцевых ветвей, nn. digit?les dors?les для V, IV пальцев и локтевой стороны III пальца.
R?mus palm?ris n. uln?ris, конечная ветвь локтевого нерва, на уровне os pisif?rme делится на поверхностную и глубокую ветви, из которых поверхностная, r?mus superfici?lis, снабжает мышечной веточкой m. palm?ris br?vis, затем кожу на локтевой стороне ладони и, разделившись, дает три nn. digit?les palm?res pr?prii к обеим сторонам мизинца и к локтевой стороне IV пальца.
R?mus prof?ndus, глубокая ветвь локтевого нерва, вместе с глубокой ветвью, a. uln?ris уходит через промежуток между m. fl?xor и m. abd?ctor d?giti m?nimi и сопровождает глубокую ладонную дугу. Там она иннервирует все мышцы hypoth?nar, все mm. inter?ssei, третью и четвертую mm. lumbric?les, а из мышц th?nar — m. add?ctor p?llicis и глубокую головку m. fl?xor p?llicis br?vis. R?mus prof?ndus оканчивается тонким анастомозом с n. medi?nus (см. рис. 310).
4. N. cut?neus br?chii medi?lis происходит из медиального пучка сплетения (из СVIII, ThI), идет по подмышечной ямке медиально от a. axill?ris, соединяется обыкновенно с прободающей ветвью II грудного нерва, так называемого n. intercostobrachi?lis, и снабжает кожу на медиальной поверхности плеча вплоть до локтевого сустава.
5. N. cut?neus antebr?chii medi?lis тоже из медиального пучка сплетения (из СVIII, ThI), в подмышечной ямке лежит рядом с n. uln?ris; в верхней части плеча располагается медиально от плечевой артерии рядом с v. bas?lica, вместе с которой прободает фасцию и становится подкожным. Нерв этот иннервирует кожу на локтевой (медиальной) стороне предплечья до лучезапястного сустава.
6. N. radi?lis, лучевой нерв (СV-СVIII, ThI), составляет продолжение заднего пучка плечевого сплетения. Он проходит сзади от плечевой артерии вместе с a. prof?nda br?chii на заднюю сторону плеча, огибает спирально плечевую кость, располагаясь в can?lis humeromuscul?ris, и затем, прободая сзади наперед латеральную межмышечную перегородку, выходит в промежуток между m. brachioradi?lis и m. brachi?lis. Здесь нерв делится на поверхностную (r?mus superfici?lis) и глубокую (r?mus prof?ndus) ветви. Перед этим n. radi?lis дает следующие ветви:
R?mi muscul?res на плече для разгибателей — m. tr?ceps и m. anc?neus. От последней веточки снабжаются еще капсула локтевого сустава и латеральный надмыщелок плеча, поэтому при воспалении последнего (эпикондилит) возникает боль по ходу всего лучевого нерва.
Nn. cut?nei br?chii post?rior et later?lis inf?rior разветвляются в коже задней и нижнем отделе заднелатеральной поверхностей плеча.
N. cut?neus anterbr?chii post?rior берет начало от лучевого нерва в can?lis humeromuscul?ris, выходит под кожу над началом m. brachioradi?lis и распространяется на тыльной стороне предплечья.
R?mi muscul?res идут к m. brachioradi?lis и m. ext?nsor c?rpi radi?lis l?ngus.
R?mus superfici?lis идет на предплечье в s?lcus radi?lis латерально от a. radi?lis, а затем в нижней трети предплечья через промежуток между лучевой костью и сухожилием m. brachioradi?lis переходит на тыл кисти и снабжает пятью тыльными ветвями, nn. digit?les dors?les, по бокам I и II палец, а также лучевую сторону III. Ветви эти обыкновенно оканчиваются на уровне последних межфаланговых суставов. Таким образом, каждый палец снабжается двумя тыльными и двумя ладонными нервами, проходящими по обеим сторонам.
Тыльные нервы происходят из n. radi?lis и n. uln?ris, иннервирующих каждый по 21/2 пальца, а ладонные — из n. medi?nus и n. uln?ris, причем первый снабжает 31/2 пальца (начиная с большого), а второй — остальные 11/2 пальца (см. рис. 311).
R?mus prof?ndus проходит сквозь m. supin?tor и, снабдив последний ветвью, выходит на дорсальную сторону предплечья, иннервируя m. ext?nsor c?rpi radi?lis br?vis и все задние мышцы предплечья. Продолжение глубокой ветви, n. inter?sseus (antebr?chii) post?rior, спускается между разгибателями большого пальца до лучезапястного сустава, который иннервирует. Из хода n. radi?lis видно, что он иннервирует все разгибатели как на плече, так и на предплечье, а на последнем еще и лучевую группу мышц. Соответственно этому на разгибательной стороне плеча и предплечья им иннервируется и кожа. Лучевой нерв — продолжение заднего пучка — является как бы задним нервом руки.
Передние ветви грудных нервов
Передние ветви, r?mi ventr?les, грудных нервов, nn. thor?cici, носят название межреберных нервов, nn. intercost?les, так как они идут в межреберных промежутках, но XII идет по нижнему краю XII ребра (n. subcost?lis) (рис. 312).
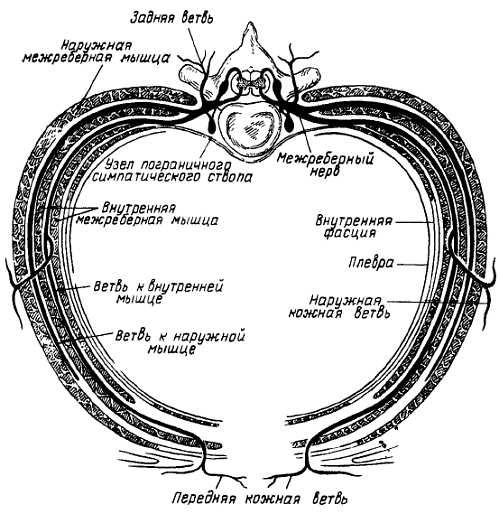
Рис. 312.
Верхние шесть межреберных нервов доходят до края грудины, нижние шесть проходят в толщу брюшной стенки, где в промежутке между поперечной и внутренней косой мышцами направляются к прямой мышце живота, куда проникают, пройдя через ее влагалище. XII межреберный нерв, проходящий по m. quadr?tus lumb?rum, близко подходит к лобковому симфизу, оканчиваясь в нижней части прямой мышцы и m. pyramid?lis.
На своем пути nn. intercost?les дают r?mi muscul?res для всех вентральных мышц в стенках грудной и брюшной полостей, а также для мышц вентрального происхождения на спине: mm. serrati posteri?res superi?res et inferi?res и mm. levat?res costarum. Они также участвуют в иннервации плевры и брюшины.
Кроме того, от nn. intercost?les отходят два ряда прободающих ветвей, снабжающих кожу на боковой поверхности груди и живота — r?mi cut?nei later?les (pector?les et abdomin?les) и на передней — r?mi cut?nei anteri?res (pector?les et abdomin?les). От них отходят ветви к молочной железе: от боковых — r?mi m?mm?rii later?les и от передних — r?mi m?mm?rii medi?les.
R?mi cut?nei anteri?res шести нижних межреберных нервов в качестве продолжения их концов прободают прямую мышцу живота и передний листок ее влагалища и разветвляются в коже живота в этой же области.
Пояснично-крестцовое сплетение
Из передних ветвей поясничных, крестцовых и копчикового нервов слагается пояснично-крестцовое сплетение, pl?xus lumbosacr?lis. Это общее сплетение разделяется по областям на частные отделы, или сплетения: поясничное, крестцовое и копчиковое.
Поясничное сплетение
Поясничное сплетение, pl?xus lumb?lis, образуется из передних ветвей трех верхних поясничных нервов и верхней части IV такого же нерва, а также веточки от XII межреберного нерва. Сплетение залегает спереди поперечных отростков поясничных позвонков в толще m. ps?as m?jor и дает целый ряд ветвей, которые выходят частью из-под латерального, частью из-под медиального края этой мышцы, частью же прободают ее и появляются на ее передней поверхности. Ветви эти следующие:
1. R?mi muscul?res к mm. ps?as m?jor et m?nor, m. quadr?tus lumb?rum и mm. intertransvers?rii later?les lumb?rum.
2. N. iliohvpog?stricus (LI) выходит из-под латерального края m. ps?as m?jor и ложится на переднюю поверхность m. quadr?tus lumb?rum параллельно XII межреберному нерву. Будучи, как и последний, сегментарным нервом, n. iliohvpog?stricus подобно ему проходит между поперечной и внутренней косой мышцами живота, снабжая их мышечными ветвями, а также иннервирует кожу верхней части ягодицы и паховою канала выше его поверхностного отверстия.
3. N. ilioinguinalis (LI) — также сегментарный нерв, выходит из-под латерального края m. ps?as m?jor и идет параллельно и книзу от n. iliohvpog?stricus, а затем непосредственно в паховом канале, выходит через поверхностное паховое кольцо и разветвляется в коже лобка и мошонки или большой половой губы.
4. N. genitofemoralis (LII) проходит сквозь толщу m. ps?as m?jor на переднюю поверхность этой мышцы и разделяется на две ветви, из которых одна, r. femor?lis, направляется к паховой связке, проходит под нее и разветвляется в коже бедра тотчас ниже этой связки. Другая ветвь, r. genit?lis, прободает заднюю стенку пахового канала и присоединяется к семенному канатику, снабжая m. crem?ster и оболочки яичка.
5. N. cut?neus f?moris later?lis (LII, LIII), выйдя из-под бокового края m. ps?as m?jor, направляется по поверхности m. ili?cus к sp?na il?aca ant?rior sup?rior, где он прободает брюшную стенку и выходит на бедро, становится подкожным и спускается по боковой поверхности бедра до колена, иннервируя кожу.
6. N. femoralis, бедренный нерв — самая толстая ветвь поясничного сплетения (LII, LIII, LIV), выходит через lac?na muscul?rum на переднюю сторону бедра. Он ложится латерально от бедренной артерии, отделяясь от нее глубоким листком, f?sciae l?tae, распадается на многочисленные ветви, из которых одни, r?mi muscul?res, иннервируют m. quadr?ceps, m. sart?rius и m. pect?neus, a другие, r?mi cut?nei anteri?res, снабжают кожу переднемедиальной поверхности бедра. Одна из кожных ветвей бедренного нерва, очень длинная, n. saph?nus, ложится в can?lis adduct?rius латерально от a. femor?lis. У hi?tus adduct?rius нерв покидает артерию, прободает переднюю стенку канала и становится поверхностным. На голени нерв сопровождает v. saph?na m?gna. От него отходит r?mus infrapatell?ris к коже нижней части колена и r?mi cut?nei cr?ris medi?les — к коже медиальной поверхности голени вплоть до такого же края стопы.
7. N. obturatorius, запирательный нерв (LII— LIV), проходит через запирательный канал на бедро и иннервирует m. obturat?rius ext?rnus, тазобедренный сустав и все приводящие мышцы вместе с m. gr?cilis и m. pect?neus, а также кожу над ними.
Крестцовое сплетение
Крестцовое сплетение, pl?xus sacr?lis, — самое значительное из всех сплетений, слагается из передних ветвей IV (нижней части) и V поясничного нерва и таких же ветвей четырех крестцовых нервов (SI—SIV), выходящих из передних отверстий крестца. Близость многочисленных пучков сплетения к крестцово-подвздошному сочленению обусловливает различные локализацию и иррадиацию болей при заболеваниях этого сочленения. Нервы сплетения, соединяясь друг с другом, образуют (рис. 313) толстый ствол седалищного нерва, выходящий через for?men infrapirif?rme из тазовой полости. Ветви, отходящие от крестцового сплетения, можно разделить на короткие и длинные.
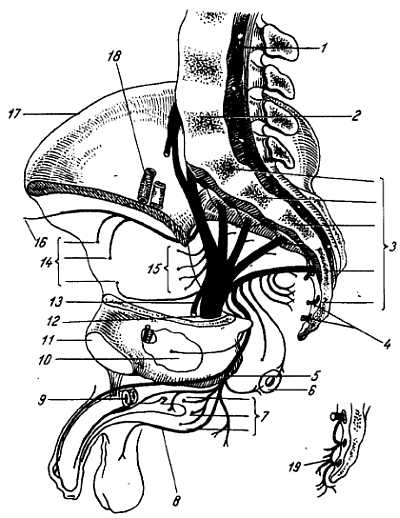
Рис. 313
1 — cauda equina; 2 — vertebra lumbalis V; 3 — vertebrae sacrales; 4 — rr. ventrales nn. sacrales; 5 — n. pudendus; 6 — ветви к rectum; 7 — ветви к мышцам промежности; 8 — ветви к половым органам: 9 — corpora cavernosa; 10 — m. obturatorius internus; 11 — facies symphysialis; 12 — n. ischiadicus; 13 — n. cutaneus femoris posterior; 14 — ветви к ягодичным мышцам; 15 — ветви к мышцам задней поверхности тазобедренного сустава; 16 — ветви к fascia lata; 17 — crista iliaca; 18 — vasa iliaca externa; 19 — plexus coccygeus.
Первые разветвляются в области пояса нижней конечности, а вторые снабжают всю нижнюю конечность, за исключением той ее части, которая снабжена ветвями поясничного сплетения.
Короткие ветви (рис. 314).
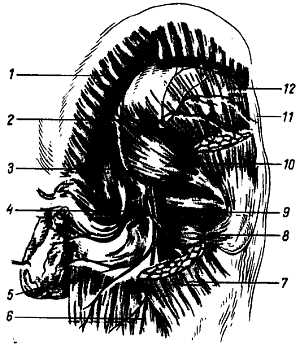
Рис. 314
1, 7 — m. gluteus maximus; 2 — n. gluteus superior; 3 — lig. sacrospinale; 4 — n. pudendus; 5 — rr. perineales; 6 — n. cutaneus femoris posterior; 8 — m. quadratus femoris; 9 — n. ischiadicus; 10 — m. piriformis; 11 — m. tensor fasciae latae; 12 — m. gluteus minimus.
1. R?mi muscul?res для m. pirif?rmis (из SI и SII), m. obturat?rius int?rnus с mm. gem?lli и quadr?tus f?moris (из LIV, LV, SI и SII), для mm. lev?tor ?ni et cocc?geus (SIII, SIV).
2. N. gl?teus sup?rior (LIV и LV и от SI) выходит через for?men suprapirif?rme из таза вместе с одноименной артерией и затем распространяется в m. gl?teus m?dius, m. gl?teus m?nimus и m. t?nsor f?sciae l?tae.
3. N. gl?teus inf?rior (LV, SI, SII), выйдя через for?men infrapirif?rme, снабжает своими ветвями m. gl?teus m?ximus и капсулу тазобедренного сустава.
4. N. pudendus (SI-SIV), выйдя через for?men infrapirif?rme, уходит обратно в таз через for?men ischi?dicum m?nus. Далее n. pud?ndus вместе с соименной артерией проходит по боковой стенке f?ssa ischiorect?lis. В пределах последней от него отходят nn. rect?les inferi?res, которые снабжают наружный сфинктер, m. sph?ncter ?ni ext?rnus, и кожу в ближайшей окружности заднего прохода. На уровне седалищного бугра у заднего края diaphr?gma urogenit?le n. pud?ndus делится на nn. perin?i и n. dors?lis p?nis (clit?ridis). Первые, идя кпереди, иннервируют m. ischiocavern?sus, m. bulbospongi?sus и m. transv?rsus perin?i superfici?lis, а также кожу промежности. Конечные ветви снабжают кожу задней стороны мошонки (nn. scrot?les posteri?res) или больших половых губ (nn. labi?les posteri?res). N. dors?lis p?nis (clit?ridis) сопровождает в толще diaphr?gma urogenit?le a. dors?lis p?nis, дает веточки к m. transv?rsus perin?i prof?ndus и m. sphincter ur?thrae, проходит на спинку полового члена (или клитора), где распространяется в коже главным образом glans p?nis. В составе n. pud?ndus проходит большое число вегетативных волокон.
Длинные ветви. 1. N. cut?neus f?moris post?rior (SI, SII, SIII) выходит из таза вместе с седалищным нервом, а затем спускается вниз под m. gl?teus m?ximus на заднюю поверхность бедра. С медиальной своей стороны он дает веточки, идущие под кожу нижней части ягодицы (nn. cl?nium inferi?res) и к промежности (r?mi perine?les). На бедре на поверхности задних мышц доходит книзу до подколенной ямки и дает многочисленные ветви, которые распространяются в коже задней стороны бедра и голени.
2. N. ischi?dicus, седалищный нерв — самый крупный из нервов всего тела, представляет непосредственное продолжение крестцового сплетения, содержащее волокна всех его корешков. Выйдя из тазовой полости через большое седалищное отверстие ниже m. pirif?rmis, прикрывается m. gl?teus m?ximus. Дальше книзу нерв выходит из-под нижнего края этой мышцы и спускается отвесно на задней стороне бедра под сгибателями голени. В верхней части подколенной ямки он обыкновенно делится на две свои главные ветви: медиальную, более толстую, n. tibi?lis, и латеральную, потоньше, n. peron?is (fibul?ris) comm?nis. Довольно часто нерв бывает разделен на два отдельных ствола уже на всем протяжении бедра.
Ветви седалищного нерва.
1. R?mi muscul?res к задним мышцам бедра: m. semitendin?sus, m. semimembran?sus и к длинной головке m. b?ceps f?moris, а также к задней части m. add?ctor m?gnus. Короткая головка m. b?ceps получает веточку от малоберцового нерва. Отсюда же отходит веточка к коленному суставу.
2. N. tibi?lis, большеберцовый нерв (LIV, LV, SI, SIII), идет прямо вниз посередине подколенной ямки по тракту подколенных сосудов, затем входит в can?lis cruropopl?teus и, сопровождая в нем а. и vv. tibi?les posteri?res, доходит до медиальной лодыжки. Позади последней n. tibi?lis разделяется на свои конечные ветви, nn. plant?res later?lis et medi?lis, проходящие в одноименных бороздках подошвы. В подколенной ямке от n. tibi?lis отходят r?mi muscul?res к m. gastrocn?mius, m. plant?ris, m. sleus и m. popl?teus, а также несколько веточек к коленному суставу. Кроме того, в подколенной ямке большеберцовый нерв отдает длинную кожную ветвь, n. cut?neus s?rae medi?lis, которая идет вниз вместе с v. saph?na p?rva и иннервирует кожу заднемедиальной поверхности голени. На голени n. tibi?lis дает n. inter?sseus cr?ris, который иннервирует все три глубокие мышцы: m. tibi?lis posterior, m. fl?xor h?llucis l?ngus и m. fl?xor digit?rum l?ngus, заднюю сторону голеностопного сустава и дает позади медиальной лодыжки кожные ветви к коже пятки и медиального края стопы (рис. 315).
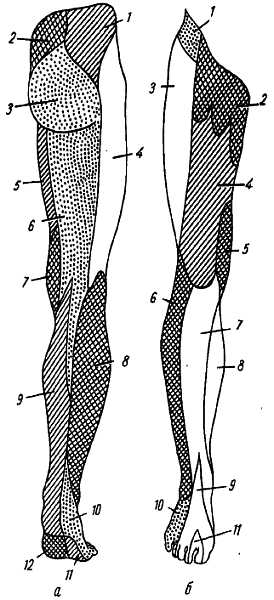
Рис. 315.
а — задняя поверхность: 1 — nn. clunium superiores; 2 — nn. clunium medii; 3 — clunium inferiores; 4 — n. cutaneus femoris lateralis; 5 — rr. cutanei anterioris n. femoralis; 6 — n. cutaneus femoris posterior; 7 — r. cutaneus n. obturatorii; 8 — n. cutaneus surae lateralis (om n. peroneus communis); 9 — n. cutaneus surae medialis (от n. tibialis); 10 — n. suralis; 11 — n. plantaris lateralis; 12 — n. plantaris medialis;
б — передняя поверхность: 1 — r. cutaneus lateralis n. iliohypogastricl; 2 — n. genitofem oralis; 3 — n. cutaneus femoris lateralis; 4 — rr. cutanei untcriores n. femoralis; 5 — r. cutancus n. obturutorii; 6 — n. cutaneus surae lateralis (or n. peroneus comm.); 7 — n. saphenus (от n. femoralis); 8 — n. cutaneus surue medialis; 9 — n. peroneus superflcialis; 10 — n. suralis; 11 — n. peroneus profundus.
N. plant?ris medi?lis вместе с одноименной артерией проходит в s?lcus plant?ris medi?lis вдоль медиального края m. fl?xor digit?rum br?vis и снабжает эту мышцу и мышцы медиальной группы, за исключением m. add?ctor h?llucis и латеральной головки m. fl?xor h?llucis br?vis. Затем нерв в конце концов распадается на семь nn. digit?les plant?res pr?prii, из которых один идет к медиальному краю большого пальца и попутно снабжает также первую и вторую mm. lumbric?les, а остальные шесть иннервируют кожу обращенных друг к другу сторон пальцев начиная с латеральной стороны большого и кончая медиальным краем IV (рис. 316).
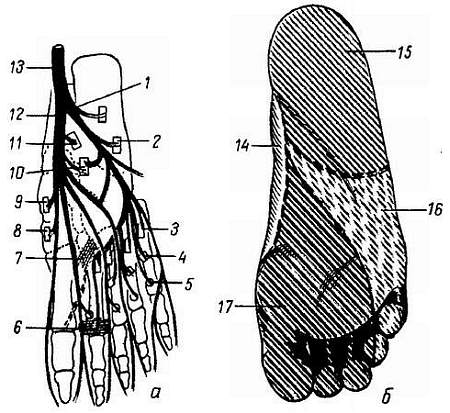
Рис. 316.
1 — n. plantaris lateralis; 2 — m. flexor digiti minimi brevis; 3 — mm. interossei dorsales; 4 — mm. interossei plantares; 5 — mm. lumbricales; 6 — caput transversum m. adductoris hallucis; 7 — caput obliquum m. adductoris hallucis; 8 — m. flexor hallucis brevis; 9 — m. abductor hallucis; 10 — m. quadratus plantae; 11 — m. flexor digitorum brevis; 12 — n. plantaris medialis; 13 — n. tibialis; 14 — область распространения ветвей n. sapheni; 15 — область иннервации n. tibialis; 16 — область иннервации n. plantaris lateralis; 17 — область иннервации n. plantaris medialis.
N. plant?ris later?lis идет по ходу одноименной артерии в s?lcus plant?ris later?lis. Иннервирует посредством r?mi muscul?res все три мышцы латеральной группы подошвы и m. quadr?tus pl?ntae и делится на две ветви — глубокую и поверхностную. Первая, r?mus prof?ndus, идет вместе с подошвенной артериальной дугой и снабжает третью и четвертую mm. lumbric?les и все mm. inter?ssei, а также m. add?ctor h?llucis и латеральную головку m. fl?xor h?llucis br?vis. Поверхностная ветвь, r?mus superfici?lis, дает ветви к коже подошвы и разделяется на три nn. digit?les plant?res pr?prii, идущие к обеим сторонам V пальца и к обращенной к последнему стороне IV пальца. В общем распределение nn. plant?res medi?lis et later?lis соответствует ходу n. medi?nus и n. uln?ris на кисти.
3. N. peron?us (fibul?ris) comm?nis, общий малоберцовый нерв (LIV, LV, SI, SII), идет латерально от n. tibi?lis к головке малоберцовой кости, где он прободает начало m. peron?us l?ngus и делится на поверхностную и глубокую ветви. На своем пути n. peron?us comm?nis дает n. cut?neus s?rae later?lis, иннервирующий кожу латеральной стороны голени. Ниже середины последней n. cutan?us s?rae later?lis соединяется с n. cut?neus s?rae medi?lis, образуя n. sur?lis, который огибает сзади латеральную лодыжку, давая ветви к коже пятки (r?mi calc?nei later?les), а затем идет под названием n. cut?neus dors?lis later?lis по латеральному краю тыла стопы, снабжая кожу этого края и боковую сторону мизинца.
Поверхностная ветвь малоберцового нерва, n. peron?us (fibul?ris) superfici?lis, спускается между mm. paron?i в can?lis musculoperon?us sup?rior, отдавая к ним мышечные ветви. На границе средней и нижней третей голени он в качестве уже только кожного нерва прободает фасцию и спускается на середину тыла стопы, разделяясь на две ветви. Одна из них, n. cut?neus dors?lis medi?lis, иннервирует медиальную сторону большого пальца и обращенные друг к другу края II и III пальцев (n?rvi digit?les dors?les). Другая ветвь, n. cut?neus dors?lis interm?dius, делится на nn. digit?les p?dis, иннервирующие обращенные друг к другу стороны тыльной поверхности II–V пальцев (см. рис. 315).
Глубокая ветвь малоберцового нерва, n. peron?us (fibul?ris) prof?ndus, проходит в сопровождении a. tibi?lis ant?rior, отдавая ветви к m. tibi?lis ant?rior, m. ext?nsor digit?rum l?ngus и m. ext?nsor h?llucis l?ngus, а также r?mus articul?ris к голеностопному суставу. N. peron?us prof?ndus вместе с сопровождающей его артерией выходит на тыл стопы, иннервирует короткий разгибатель пальцев и затем, разделившись на два nn. digit?les dors?les, снабжает кожу обращенных друг к другу поверхностей I и II пальцев.
В составе крестцового сплетения, относящегося к анимальной нервной системе, проходят преганглионарные, парасимпатические волокна, начинающиеся в боковых рогах II–IV крестцовых сегментов спинного мозга. Эти волокна в виде n?rvi spl?nchnici pelv?ni направляются к нервным сплетениям таза, иннервирующим тазовые внутренности: мочевой пузырь, сигмовидную и прямую кишку и внутренние половые органы.
Копчиковое сплетение
Копчиковое сплетение, pl?xus cocc?geus, составляется передними ветвями V крестцового и копчикового нервов. Из него исходят тонкие nn. anococc?gei, которые, соединившись с задней ветвью копчикового нерва, разветвляются в коже у верхушки копчика.
ЧЕРЕПНЫЕ НЕРВЫ
Черепных нервов, nn. crani?les (encephalici), 12 пар: I — nn. olfact?rii, II — n. ?pticus, III — n. oculomot?rius, IV — n. trochle?ris, V — n. trig?minus, VI — n. abd?cens, VII — n. faci?lis, VIII — n. vestibulocochle?ris, IX — n. glossophar?ngeus, X — n. v?gus, XI — n. access?rius, XII — n. hypogl?ssus.
Черепные нервы имеют особенности, отличающие их от спинномозговых нервов. Эти особенности зависят главным образом от иных условий развития мозга и головы сравнительно со спинным мозгом и туловищем. Прежде всего первые два черепных нерва, связанные с передним мозгом, по своему характеру и происхождению занимают совершенно отдельное положение среди всех нервов. Они являются выростами мозга. Остальные черепные нервы, хотя принципиально и не отличаются от спинномозговых нервов, но тем не менее для них характерно то обстоятельство, что ни один из них не соответствует полному спинномозговому нерву, слагающемуся из переднего и заднего корешков. Каждый из черепных нервов представляет собой какой-нибудь один из этих двух корешков, которые в области головы никогда не соединяются вместе, что напоминает подобные же отношения, существующие у спинномозговых нервов примитивных позвоночных (миноги) (рис. 317). III, IV, VI, XI и XII черепные нервы соответствуют передним корешкам спинномозговых нервов, а V, VII, VIII, IX и X нервы гомологичны задним (рис. 318, 319).
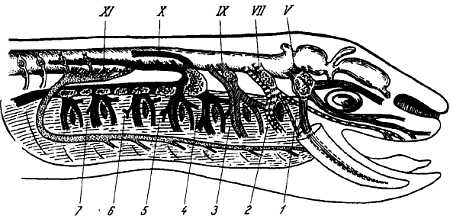
Рис. 317
Рис. 318.
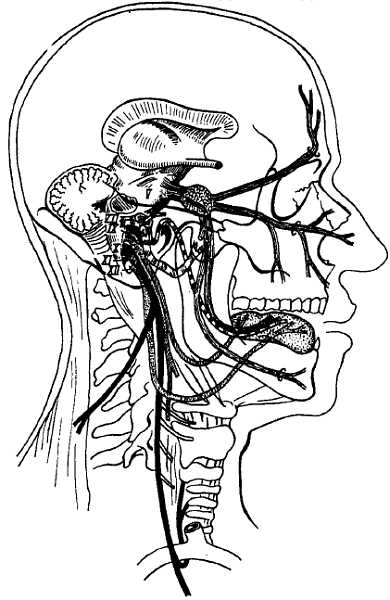
Рис. 319.
Особенности черепных нервов связаны с прогрессивным развитием головного мозга.
Черепные нервы, как и спинномозговые, имеют ядра серого вещества: соматически-чувствительные (соответствующие задним рогам серого вещества спинного мозга), соматически-двигательные (соответствующие передним рогам) и вегетативные (соответствующие боковым рогам). Последние можно разделить на висцерально-чувствительные и висцерально-двигательные, из которых висцерально-двигательные иннервируют не только неисчерченную (гладкую) мускулатуру, но и скелетные мышцы висцерального происхождения. Учитывая, что исчерченные (скелетные) мышцы приобрели черты соматических мускулов, все ядра черепных нервов, имеющих отношение к таким мышцам независимо от их происхождения, лучше обозначать как соматически-двигательные.
В результате в составе черепных нервов имеются те же компоненты, что и в спинномозговых нервах.
Афферентные:
1. Соматически-чувствительные волокна, идущие от органов, воспринимающих физические раздражители (давление, температуру, звук и свет), т. е. от кожи, органов слуха и зрения, — II, V, VIII.
2. Висцерально-чувствительные волокна, идущие от органов, воспринимающих химические раздражители (растворенные или взвешенные в окружающей среде или во внутренних полостях частицы различных веществ), т. е. от нервных окончаний в органах пищеварения и других внутренностях, от специальных органов глотки, ротовой (органы вкуса) и носовой (органы обоняния) полостей, — I, V, VII, IX, X.
Эфферентные:
3. Соматически-двигательные волокна, иннервирующие произвольную мускулатуру, а именно: мышцы, происшедшие из головных миотомов, глазные мышцы (III, IV, VI), и подъязычную мускулатуру (XII), а также вторично сместившиеся в состав переднего отдела пищеварительного тракта мышцы скелетного типа — так называемые мышцы жаберного аппарата, ставшие у млекопитающих и человека жевательными, мимическими и т. п. (V, VII, IX, X, XI).
4. Висцерально-двигательные волокна, иннервирующие висцеральную мускулатуру, т. е. непроизвольную мускулатуру сосудов и внутренностей (органы пищеварения и дыхания), мышцу сердца, а также различного рода железы (секреторные волокна), — VII, IX, X.
В составе двигательных нервов к тем же органам проходят симпатические волокна, идущие из соответствующих симпатических узлов.
Из 12 пар черепных нервов соматически-чувствительным является VIII нерв, соматически-двигательными — III, IV, VI, XI, XII. Остальные нервы (V, VII, IX, X) являются смешанными.
Обонятельный нерв, который можно назвать висцерально-чувствительным, и зрительный — соматически-чувствительный занимают особое положение, что уже отмечалось.
Малое число соматически-двигательных нервов по сравнению с остальными находится в связи с редукцией миотомов головы, дающих начало лишь глазным мышцам. Развитие смешанных нервов, содержащих висцеральные компоненты, связано с эволюцией передней части кишечной трубки (хватательной и дыхательной), в области которой развивается висцеральный аппарат со сложной чувствующей областью и значительной мускулатурой.
Нервы, развившиеся путем слияния спинномозговых нервов
К этой группе относится один нерв — n. hypogl?ssus.
Подъязычный нерв (XII)
N. hypogl?ssus, подъязычный нерв, есть результат слияния 3–4 спинномозговых (затылочных) сегментарных нервов, существующих у животных самостоятельно и иннервирующих подъязычную мускулатуру. Соответственно обособлению из нее мышц языка эти нервы (затылочные и передние спинномозговые) у высших позвоночных и человека сливаются вместе, образуя как бы переходную группу от спинномозговых нервов к черепным. Этим объясняются положение ядра нерва не только в головном мозге, но и в спинном, положение самого нерва в переднелатеральной борозде продолговатого мозга вблизи спинного мозга и выход его многими корешковыми нитями (10–15), а также связь с передними ветвями I и II шейных нервов в виде ?nsa cervic?lis (см. рис. 272; рис. 320).
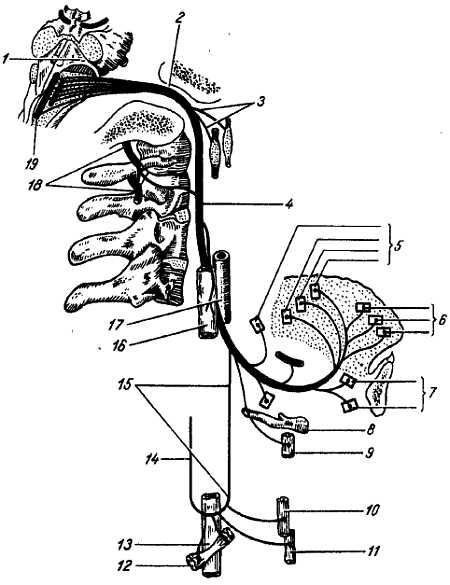
Рис. 320.
1 — fossa rhomboidea; 2 — canalis hypoglossalis; 3 — соединительные ветви подъязычного нерва с верхним симпатическим шейным узлом и с нижним узлом блуждающего нерва; 4 — n. hypoglossus; 5, 6 — ветви подъязычного нерва к мышцам языка; 7 — ветви к m. geniohyoideus; 8 — os hyoideum; 9 — ветвь к щитоподъязычной мышце; 10 — ветви к грудино-подъязычной мышце; 11 — ветви к грудино-щитовидной мышце; 12 — ветвь к лопаточно-подъязычной мышце; 13 — внутренняя яремная вена; 14 — нижний корешок шейной петли; 15 — верхний корешок шейной петли; 16 — внутренняя яремная вена; 17 — внутренняя сонная артерия: 18 — nn. cervicales; 19 — nucl. n. hypoglossi.
Подъязычный нерв, являясь мышечным, содержит эфферентные (двигательные) волокна к мышцам языка и афферентные (проприоцептивные) волокна от рецепторов этих мышц. В нем проходят также симпатические волокна от верхнего шейного симпатического узла. Он имеет связи с n. lingu?lis, с нижним узлом n. v?gi, с I и II шейными нервами.
Единственное соматически-двигательное ядро нерва, заложенное в продолговатом мозге, в области trig?num n. hypogl?ssi ромбовидной ямки, спускается через продолговатый мозг, доходя до I–II шейного сегмента; оно входит в систему ретикулярной формации. Появляясь на основании мозга между пирамидой и оливой несколькими корешками, нерв затем проходит через одноименный канал затылочной кости, can?lis hypogl?ssalis, спускается по латеральной стороне a. car?tis int?rna, проходит под задним брюшком m. dig?stricus и идет в виде дуги, выпуклой книзу, по латеральной поверхности m. hyogl?ssus. Здесь дуга подъязычного нерва ограничивает сверху треугольник Пирогова.
При высоком расположении дуги подъязычного нерва треугольник Пирогова имеет большую площадь и наоборот. У переднего края m. hyogl?ssus подъязычный нерв распадается на свои конечные ветви, которые входят в мускулатуру языка. Часть волокон подъязычного нерва идет в составе ветвей лицевого нерва к круговой мышце рта, почему при поражении ядра нерва несколько страдает и функция этой мышцы.
Одна из ветвей нерва, r?dix sup?rior, спускается вниз, соединяется с r?dix inf?rior шейного сплетения и образует вместе с ним шейную петлю — ?nsa cervic?lis (рис. 321).
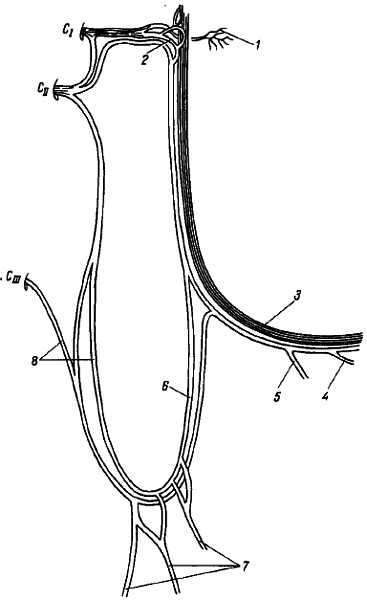
Рис. 321.
1 — ветви к m. rectus capitis anterior и к m. longus capitis; 2 — пучок волокон из С1 в составе radix superior n. hypoglossi; 3 — n. hypoglossus; 4 — ветвь к m. geniohyoideus; 5 — ветвь к m. thyrohyoideus; 6 — radix superior n. hypoglossi; 7 — rr. musculares; 8 — radix inferior шейного сплетения в составе шейной петли.
Следовательно, ?nsa cervic?lis — шейная петля, представляет соединение последнего черепного нерва (подъязычного) с первым сплетением спинномозговых нервов, шейным сплетением. От этой петли иннервируются мышцы, расположенные ниже подъязычной кости, и m. geniohyo?deus. R?dix sup?rior подъязычного нерва состоит целиком из волокон I и II шейных нервов, присоединившихся к нему из шейного сплетения.
Эту морфологическую связь подъязычного нерва с шейным сплетением можно объяснить развитием нерва, а также тем, что мышцы языка при акте глотания функционально тесно связаны с мышцами шеи, действующими на подъязычную кость и щитовидный хрящ.
Нервы жаберных дуг
К этой группе относятся V, VII, IX и X черепные нервы, которые как гомологи задних корешков спинномозговых нервов снабжены лежащими вне мозга нервными узлами с находящимися в них псевдоуниполярными клетками. Эти нервы развиваются в связи с задним (ромбовидным) мозгом. Наряду с чувствительными волокнами они содержат в себе и двигательные, иннервирующие мускулатуру жаберного аппарата.
Типичный висцеральный нерв у рыб, обслуживающий висцеральную (жаберную) дугу, обычно состоит из наджаберного узла, g?nglion epibranchi?le (от греч. branchiae — жабры), преджаберной ветви, r?mus pretrem?ticus, состоящей из чувствительных волокон, и зажаберной ветви, r?mus posttrem?ticus, содержащей и чувствительные, и двигательные волокна. Чувствительные волокна обеих ветвей являются отростками нейронов, лежащих в наджаберном узле, а двигательные проходят мимо узла, как в спинномозговом нерве. Эти характерные черты строения типичного висцерального нерва и будут проявляться более или менее отчетливо в строении указанных нервов.
В этой группе будут описаны также XI пара, n. access?rius, который является отщеплением X нерва, и VIII пара, n. vestibulocochle?ris. Последний является афферентным нервом, обособившимся в процессе развития от лицевого нерва, и поэтому, хотя он и не относится к нервам жаберных дуг, данные о нем будут изложены после VII пары.
Тройничный нерв (V)
N. trig?minus, тройничный нерв, развивается в связи с первой жаберной дугой (мандибулярной) и является смешанным. Чувствительными своими волокнами иннервирует кожу лица и передней части головы, граничит сзади с областью распространения в коже задних ветвей шейных нервов и ветвей шейного сплетения. Кожные ветви (задние) II шейного нерва заходят на территорию тройничного нерва, вследствие чего возникает пограничная зона смешанной иннервации шириной в 1–2 поперечника пальца. Тройничный нерв также является проводником чувствительности от рецепторов слизистых оболочек рта, носа, уха и конъюнктивы глаза, кроме тех отделов их, которые являются специфическими рецепторами органов чувств (иннервируемых из I, II, VII, VIII и IX пар) (см. рис. 272; рис. 322).
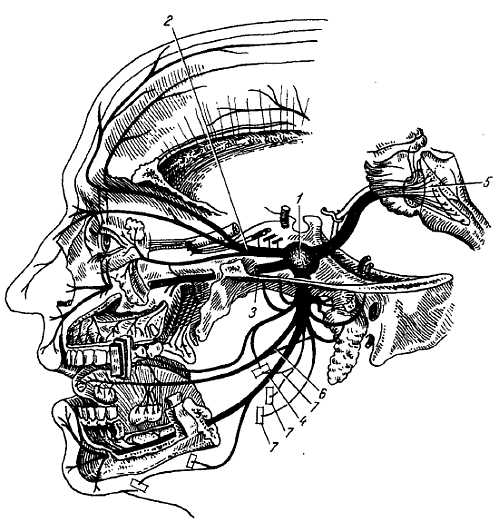
Рис. 322.
1 — gangl. trigeminale; 2 — первая ветвь n. trigemini; 3 — вторая ветвь n. trigemini; 4 — третья ветвь n. trigemini; 5 — дно IV желудочка; 6 — n. lingualis; 7 — ветви к жевательной мускулатуре.
В качестве нерва первой жаберной дуги n. trig?minus иннервирует развившиеся из нее жевательные мышцы и мышцы дна полости рта и содержит исходящие от их рецепторов афферентные (проприоцептивные) волокна, заканчивающиеся в n?cleus mesenceph?licus n. trig?mini.
В составе ветвей нерва проходят, кроме того, секреторные (вегетативные) волокна к железам, находящимся в области лицевых полостей.
Поскольку тройничный нерв является смешанным, он имеет четыре ядра, из которых два чувствительных и одно двигательное заложены в заднем мозге, а одно чувствительное (проприоцептивное) — в среднем мозге. Отростки клеток, заложенных в двигательном ядре (n?cleus mot?rius), выходят из моста на линии, отделяющей мост от средней ножки мозжечка и соединяющей место выхода nn. trig?mini et faci?lis (l?nea trigeminofaci?lis), образуя двигательный корешок нерва, r?dix mot?ria. Рядом с ним в вещество мозга входит чувствительный корешок, r?dix sens?ria. Оба корешка составляют ствол тройничного нерва, который по выходе из мозга проникает под твердую оболочку дна средней черепной ямки и ложится на верхнюю поверхность пирамиды височной кости у ее верхушки, там, где находится impr?ssio trig?mini. Здесь твердая оболочка, раздваиваясь, образует для него небольшую полость, c?vum trigemin?le. В этой полости чувствительный корешок имеет большой тройничный узел, g?nglion trigemin?le. Центральные отростки клеток этого узла составляют r?dix sens?ria и идут к чувствительным ядрам: n?cleus pontinus n. trig?mini, n?cleus spin?lis n. trig?mini и n?cleus mesenceph?licus n. trig?mini, а периферические идут в составе трех главных ветвей тройничного нерва, отходящих от выпуклого края узла.
Ветви эти следующие: первая, или глазная, n. ophth?lmicus, вторая, или верхнечелюстная, n. maxill?ris, и третья, или нижнечелюстная, n. mandibul?ris. Двигательный корешок тройничного нерва, не принимающий участия в образовании узла, проходит свободно под последним и затем присоединяется к третьей ветви. Тройничный нерв человека является результатом слияния двух нервов животных: 1) n. ophth?lmicus prof?ndus, или n. trig?minus I, и 2) n. maxillomandibul?ris, или n. trig?minus II. Следы этого слияния бывают заметны и в g?nglion trigemin?le нерва, который часто бывает двойным. Соответственно этому r?mus ophth?lmicus есть бывший n. ophth?lmicus prof?ndus, а две остальные ветви составляют n. maxillomandibul?ris, который, являясь нервом первой жаберной дуги, имеет строение типичного висцерального нерва: g?nglion trig?minale его гомологичен наджаберному узлу, r?mus maxill?ris — преджаберной ветви, a r?mus mandibul?ris — зажаберной ветви. Этим объясняется, что r?mus mandibul?ris является смешанной ветвью, а r?dix mot?ria минует узел нерва.
Каждая из трех ветвей тройничного нерва посылает тонкую веточку к твердой оболочке головного мозга.
В области разветвлений каждой из трех ветвей n. trig?minus находится еще несколько небольших нервных узелков, относящихся к вегетативной нервной системе, но описываемых обыкновенно при тройничном нерве. Эти вегетативные (парасимпатические) узлы образовались из клеток, выселившихся в процессе эмбриогенеза по путям ветвей тройничного нерва, чем и объясняется сохранившаяся на всю жизнь связь с ними, а именно: с n. ophth?lmicus — g?nglion cili?re, с n. maxill?ris — g. pterygopalat?num, с n. mandibul?ris — g. ?ticum и с n. lingu?lis (из третьей ветви) — g. submandibul?re.
Первая ветвь тройничного нерва (рис. 323). N. ophth?lmicus, глазной нерв, выходит из полости черепа в глазницу через fiss?ra orbit?lis sup?rior, но перед вступлением в нее еще делится на три ветви: n. front?lis, n. lacrim?lis и n. nasocili?ris.
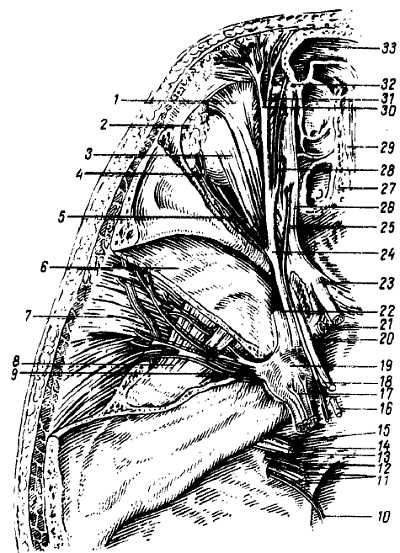
Рис. 323
1 — m. levator palpebrae superioris; 2 — gl. lacrimalis; 3 — m. rectus oculi superior; 4 — n. lacrimalis; 5 — m. rectus oculi lateralis; 6 — fossa cranii media; 7 — m. temporalis; 8 — m. pterygoideus lateralis; 9 — n. mandibuluris; 10 — n. accessorius; 11 — n. vagus; 12 — n. glossophuryngeus; 13 — n. cochlearis VIII пары; 14 — p. vestibularis VIII пары; 15 — n. facialis; 16, 18 — n. nbducens; 17 — n. trigeminus; 19 — gangl. trigeminale; 20 — n. oculomotorius; 21 — a. carmis interna; 22 — n. maxillaris; 23 — n. opticus; 24 — n. ophthalmicus; 25 — n. trochlcarb; 26 — m. obliquus oculi suporior; 27 — lam. cribrosa; 28 — n. nasoeiliaris; 29 — crista galli; 30 — n. supraorbitulis; 31 — n. frontalis; 32 — trochlea; 33 — sinus frontalis.
1. N. front?lis, лобный нерв, направляется прямо кпереди под крышей глазницы через incis?ra (или for?men) supraorbit?lis в кожу лба, здесь он называется n. supraorbit?lis, давая по пути ветви в кожу верхнего века и медиального угла глаза.
2. N. lacrim?lis, слезный нерв, идет к слезной железе и, пройдя через нее, оканчивается в коже и конъюнктиве латерального угла глаза. До входа в слезную железу n. lacrim?lis соединяется с n. zygom?ticus (от второй ветви тройничного нерва). Через этот «анастомоз» n. lacrim?lis получает секреторные волокна для слезной железы и снабжает ее также чувствительными волокнами.
3. N. nasocili?ris, носоресничный нерв, иннервирует переднюю часть носовой полости (nn. ethmoid?les ant?rior et post?rior), глазное яблоко (nn. cili?res l?ngi), кожу медиального угла глаза, конъюнктиву и слезный мешок (n. infratrochlearis). От него отходит также соединительная ветвь к g?nglion cili?re. N. ophth?lmicus осуществляет чувствительную (проприоцептивную) иннервацию глазных мышц при помощи связей с III, IV и VI нервами.
G?ndlion cili?re, ресничный узел, в форме продолговатого комочка около 1,5 мм длиной лежит в задней части глазницы на боковой стороне зрительного нерва. В этом узле, относящемся к вегетативной нервной системе, прерываются парасимпатические волокна, идущие из добавочного ядра глазодвигательного нерва в составе n. oculomot?rius к мышцам глаза. От переднего конца узла отходят 3–6 nn. cili?res br?ves, которые прободают склеру глазного яблока в окружности зрительного нерва и идут внутрь глаза. Через эта нервы проходят (после перерыва их в узле) указанные парасимпатические волокна к m. sph?ncter pup?llae и m. cili?ris.
Вторая ветвь тройничного нерва (рис. 324).
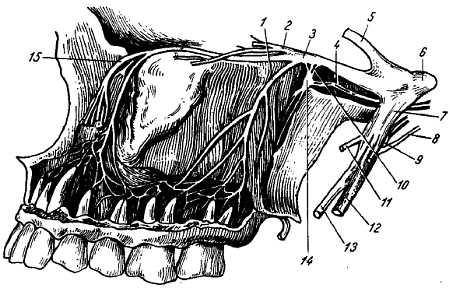
Рис. 324
1 — r. dentalis superior; 2 — n. zygomaticus; 3 — n. maxillaris; 4 — n. canalis pterygoidei; 5 — n. ophthalmicus; 6 — n. trigeminus; 7 — n. mandibularis; 8 — chorda tympani; 9 — gangl., oticum; 10 — веточки gangl. pterygopalatinum к n. maxillaris; 11 — n. massetericus; 12 — n. alveolaris inferior; 13 — n. lingualis; 14 — gangl. pterygopalatinum; 15 — n. infraorbitalis.
N. maxill?ris, верхнечелюстной нерв, выходит из полости черепа через for?men rot?ndum в крыловидно-небную ямку; отсюда его непосредственным продолжением является n. infraorbit?lis, идущий через fiss?ra orbit?lis inf?rior в s?lcus и can?lis infraorbit?lis на нижней стенке глазницы и затем выходящий через for?men infraorbit?le на лицо[36], где он распадается на пучок ветвей. Ветви эти, соединяясь отчасти с ветвями n. faci?lis, иннервируют кожу нижнего века, боковой поверхности носа и верхней губы.
От n. maxill?ris и его продолжения, n. infraorbit?lis, отходят, кроме того, следующие ветви:
1. N. zygom?ticus, скуловой нерв, к коже щеки и передней части височной области.
2. Nn. alveol?res superi?res в толще max?lla образуют сплетение, pl?xus derit?lis sup?rior, от которого отходят r?mi dent?les superi?res к верхним зубам и r?mi gingiv?les superi?res к деснам.
3. Rr. ganglion?res соединяют n. maxill?ris с g?nglion pterygopalat?num.
G?nglion pterygopalat?num, крылонебный узел, расположен в крыловидно-небной ямке медиально и книзу от n. maxillaris. В узле, относящемся к вегетативной нервной системе, прерываются парасимпатические волокна, идущие из вегетативного ядра n. interm?dius к слезной железе и железам оболочки слизистой носа и неба в составе самого нерва и далее в виде n. petr?sus major (ветвь лицевого нерва).
G?nglion pterygopalat?num отдает следующие (секреторные) ветви (рис. 325):
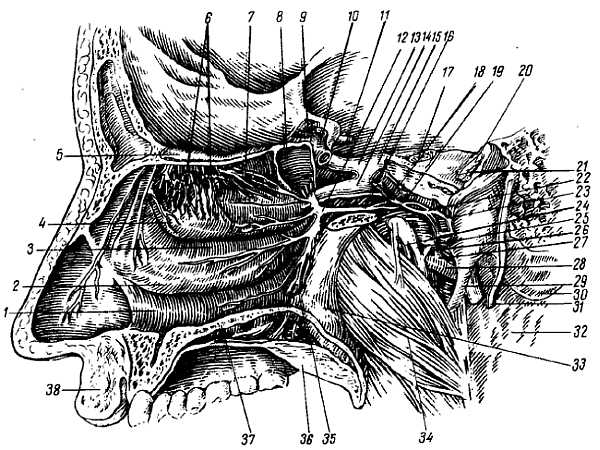
Рис. 325.
1 — meatus nasi inferior; 2, 4, 7 — conchae nasales inferior, media et superior; 3 — meatus nasi medtus; 5 — bulbus olfactorius; 6 — nn. olfactorii; 8 — sinus sphenoidalis; 9 — n. opticus; 10, 23 — a. carotis interna; 11 — n. oculomotorius; 12 — gangl. pterygopalatinum; 13 — n. ophthalmicus; 14 — n. maxillaris; 15 — gangl. trigeminale; 16 — n. canalis pterygoidei; 17 — n. trigeminus; 18 — n. petrosus major; 19 — n. petrosus profundus; 20, 31 — n. facialis; 21 — VIII пара черепных нервов; 22 — plexus sympathicus вокруг a. carotis interna; 24 — n. lingualis; 25 — n. alveolaris inferior; 26 — chorda tympani; 27 — a. meningea media; 28 — a. maxillaris; 29 — processus styloideus; 30 — processus mastoideus; 32 — gI. parotis; 33 — lam. perpendicularis ossis palatini; 34 — m. pterygoideus medialis; 35 — nn. palatini; 36 — palatum molle; 37 — palatum durum; 38 — labium superius.
1) r?mi nas?les posteri?res идут через for?men sphenopalat?num к железам слизистой оболочки носа; наиболее крупная из них, n. nasopalatinus, проходит через, can?lis incis?vus, к железам слизистой оболочки твердого неба; 2) nn. palat?ni спускаются по can?lis palat?nus m?jor и, выходя через for?mina palat?na m?jus et m?nus, иннервируют железы слизистой оболочки твердого и мягкого неба.
В составе нервов, отходящих от крылонебного узла, проходят, кроме секреторных волокон, еще чувствительные (от второй ветви тройничного нерва) и симпатические волокна. Таким образом, волокна n. interm?dius (парасимпатической части лицевого нерва), проходящие по n. petr?sus m?jor, через крылонебный узел иннервируют железы носовой полости и нёба, а также слезную железу. Эти волокна идут из крылонебного узла через n. zygom?ticus, а из него в n. lacrim?lis.
Третья ветвь тройничного нерва (рис. 326).
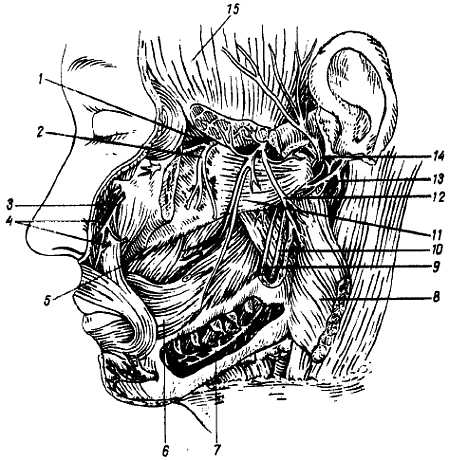
Рис. 326.
1 — n. maxillaris; 2 — n. alveolaris superior; 3, 4 — n. infraorbitalis; 5 — n. buccalis; 6 — m. buccinator; 7, 10 — n. alveolaris inferior; 8 — m. massoter; 9 — n. lingualis; 11 — m. pterygoideus laterulis; 12 — n. massetericus; 13 — n. facialis; 14 — n. auriculotemporalis; 15 — m. temporalis.
N. mandibul?ris, нижнечелюстной нерв, имеет в своем составе, кроме чувствительного, весь двигательный корешок тройничного нерва, идущий из упомянутого двигательного ядра, n?cleus mot?rius, к мускулатуре, возникшей из нижнечелюстной дуги, а потому иннервирует мышцы, прикрепляющиеся к нижней челюсти, кожу, ее покрывающую, и другие производные нижнечелюстной дуги. По выходе из черепа через for?men ov?le он делится на две группы ветвей.
А. Мышечные ветви:
К соименным мышцам: n. masset?ricus, nn. tempor?les prof?ndi, nn. pterygo?dei medi?lis et later?lis, n. tens?ris t?mpani, n. tens?ris v?li palat?ni, n. mylohyo?deus; последний отходит от n. alveol?ris inf?rior, ветви n. mandibul?ris, и иннервирует также переднее брюшко m. digastricus.
Б. Чувствительные ветви:
1. N. bucc?lis к слизистой оболочке щеки.
2. N. lingu?lis ложится под слизистую оболочку дна полости рта. Отдав n. sublingu?lis к слизистой оболочке дна полости рта, он иннервирует слизистую оболочку спинки языка на протяжении ее передних двух третей. В том месте, где n. lingu?lis проходит между обеими крыловидными мышцами, к нему присоединяется выходящая из fiss?ra petrotymp?nica тонкая веточка лицевого нерва — ch?rda t?mpani. В ней проходят исходящие из n?cleus salivat?rius sup?rior n. interm?dii парасимпатические секреторные волокна для подъязычной и поднижнечелюстной слюнных желез. Она несет также в своем составе вкусовые волокна от передних двух третей языка. Волокна самого n. lingu?lis, распространяющиеся в языке, являются проводниками общей чувствительности (осязания, боли, температурной чувствительности).
3. N. alveol?ris inf?rior через for?men mand?bulae вместе с одноименной артерией уходит в канал нижней челюсти, где дает ветви ко всем нижним зубам, предварительно образовав сплетение, pl?xus dent?lis inf?rior. У переднего конца can?lis mand?bulae n. alveol?ris inf?rior дает толстую ветвь, n. ment?lis, которая выходит из for?men ment?le и распространяется в коже подбородка и нижней губы. N. alveol?ris inf?rior — чувствительный нерв с небольшой примесью двигательных волокон, которые выходят из него у for?men mand?bulae в составе n. mylohyo?deus (см. выше).
4. N. auriculotempor?lis проникает в верхнюю часть околоушной железы и идет в височную область, сопровождая a. tempor?lis superfici?lis. Дает секреторные ветви к околоушной слюнной железе (о происхождении их см. ниже), а также чувствительные ветви к височно-нижнечелюстному суставу, к коже передней части ушной раковины, наружного слухового прохода и к коже виска.
В области третьей ветви тройничного нерва имеются два узелка, относящихся к вегетативной системе, через посредство которых происходит главным образом иннервация слюнных желез. Один из них — g?nglion ?ticum, ушной узел представляет небольшое кругловатое тело, расположенное под for?men ov?le на медиальной стороне n. mandibul?ris. К нему приходят парасимпатические секреторные волокна в составе n. petr?sus minor, являющегося продолжением n. tymp?nicus, происходящего из языкоглоточного нерва. Волокна эти прерываются в узле и идут к околоушной железе через посредство n. auriculotempor?lis, с которым g?nglion ?ticum находится в соединении. Другой узелок, g?nglion submandibul?re, поднижнечелюстной узел, располагается у переднего края m. pterygo?deus medi?lis, поверх поднижнечелюстной слюнной железы, под n. lingu?lis. Узел связан ветвями с n. lingu?lis. Через посредство этих ветвей идут к узлу и оканчиваются в нем волокна ch?rda t?mpani; продолжением их служат исходящие из g?nglion submandibul?re волокна, иннервирующие поднижнечелюстную и подъязычную слюнные железы.
Лицевой нерв (VII)
N. faci?lis (n. intermedio-facialis), лицевой нерв, является смешанным нервом; в качестве нерва второй жаберной дуги иннервирует развившиеся из нее мышцы все мимические и часть подъязычных и содержит исходящие из его двигательного ядра эфферентные (двигательные) волокна к этим мышцам и исходящие от рецепторов последних афферентные (проприоцептивные) волокна. В его составе проходят также вкусовые (афферентные) и секреторные (эфферентные) волокна, принадлежащие так называемому промежуточному нерву, n. interm?dius (см. ниже) (рис. 327).
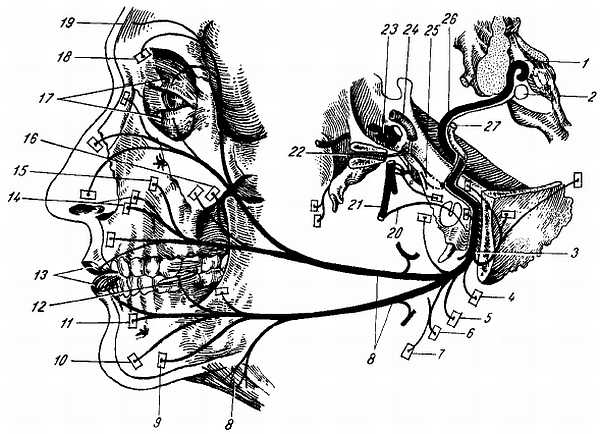
Рис. 327.
1 — дно IV желудочка; 2 — nucl. n. facialis; 3 — stylomastoideum; 4 — m. auricularis posterior; 5 — venter occipitalis; 6 — venter posterior m. digastrici; 7 — m. styiohyoideus; 8 — ветви n. facialis к мимической мускулатуре и к m. piatysma; 9 — m. depressor angulioris; 10 — m. mentalis; 11 — m. depressor labii inferioris; 12 — m. buccinator; 13 — orbicularis oris; 14 — m. levator labii superiores; 15 — levator anguli oris; 16 — zygomaticus; 17 — m. orbicularis oculi; 18 — m. corrugator supercilii; 19 — venter frontalis m. epicranii; 20 — chorda tympani; 21 — n. lingualis; 22 — gangl. pterygopalatinum; 23 — gangl. trigemjnale; 24 — carotis interna; 25 — n. intermedius; 26 — n. facialis; 27 — vestibulocochlearis.
Соответственно компонентам, составляющим его, n. faci?lis имеет три ядра, заложенных в мосту: двигательное — n?cleus mot?rius n?rvi faci?lis чувствительное — n?cleus solit?rius и секреторное — n?cleus salivat?rius sup?rior. Последние два ядра принадлежат n?rvus interm?dius.
N. faci?lis выходит на поверхность мозга сбоку по заднему краю моста, на linea trigeminofaci?lis, рядом с n. vestibulocochle?ris. Затем он вместе с последним нервом проникает в p?rus ac?sticus int?rnus и вступает в лицевой канал (can?lis faci?lis). В канале нерв вначале идет горизонтально, направляясь кнаружи, затем в области hi?tus can?lis n. petr?si maj?ris он поворачивает под прямым углом назад и также горизонтально проходит по внутренней стенке барабанной полости в верхней ее части. Миновав пределы барабанной полости, нерв снова делает изгиб и спускается вертикально вниз, выходя из черепа через for?men stylomasto?deum. В том месте, где нерв поворачивая назад, образует угол (коленце, gen?culum), чувствительная (вкусовая) часть его образует небольшой нервный узелок, g?nglion gen?culi (узел коленца).
При выходе из for?men stylomasto?deum лицевой нерв вступает в толщу околоушной железы и разделяется на свои конечные ветви. На пути в одноименном канале височной кости n. faci?lis дает следующие ветви (рис. 328):
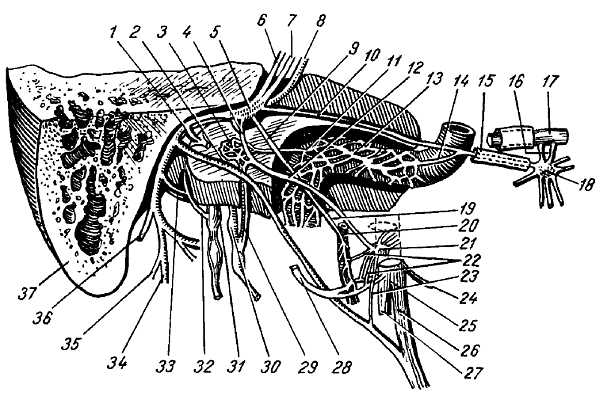
Рис. 328.
1 — n. stapedius; 2 — chorda tympani; 3 — plexus tympanicus; 4 — r. communicans n. facialis к plexus tympanicus; 5 — gangl. geniculi; 6 — n. facialis; 7 — n. intermedius; 8 — VIII пара черепных нервов; 9, 19 — r. comnunicans со сплетением вокруг a. meningea media; 10 — n. petrosus major; 11 — caroticotympanicus; 12 — n. petrosus minor; 13 — plexus sympathicus a. carotis internae; 14 — n. petros profundus; 15 — n. canalis pterygoidei; 16 — nn. pierygopalatini; 17 — n. maxillaris; 18 — gangl. pterygopalatinum; 20 — plexus sympathicus a. meningeae mediae; 21 — gangl. oticum; 22 — ветви от gangl. oticum к auriculotemporalis; 23 — r. communicans между gangl. oticum и chorda tympani; 24 — n. massetericus; 25 — n. mandibularis; 26 — n. lingualis; 27 — n. alveolaris inferior; 28 — n. auriculotemporalis; 29 — a. tympanica; 30 — n. glossopharyngeus; 31 — n. vagus (gangl. superius); 32 — r. auricularis n. vagi; 33 — r. communicans n.facialis к r. auricrularis n. vagi; 34 — ветвь n. facialis к m. styiohyoideus; 35 — ветвь n. facialis к vender posterior m. digastrici; 36 — n. auricularis posterior; 37 — processus masioideus.
1. N. petr?sus m?jor (секреторный нерв) берет начало в области коленца и выходит через hi?tus can?lis n. petr?si maj?ris; затем он направляется по одноименной бороздке на передней поверхности пирамиды височной кости, s?lcus n. petr?si maj?ris, проходит в can?lis pterygo?deus вместе с симпатическим нервом, n. petr?sus prof?ndus, образуя с ним общий n. can?lis pterygo?dei, и достигает g?nglion pterygopalat?num. Нерв прерывается в узле и его волокна в составе rami nas?les posteri?res и nn. palat?ni идут к железам слизистой оболочки носа и нёба; часть волокон в составе n. zygom?ticus (из n. maxill?ris) через связи с n. lacrim?lis достигает слезной железы.
2. N. stap?dius (мышечный) иннервирует m. stap?dius.
3. Ch?rda t?mpani (смешанная ветвь), отделившись от лицевого нерва в нижней части лицевого канала, проникает в барабанную полость, ложится там на медиальную поверхность барабанной перепонки, а затем уходит через fiss?ra petrotymp?nica. Выйдя из щели наружу, она спускается вниз и кпереди и присоединяется к n. lingu?lis.
Чувствительная (вкусовая) часть ch?rdae tympani (периферические отростки клеток, лежащих в g?nglion geniculi) идет в составе n. lingu?lis к слизистой оболочке языка, снабжая вкусовыми волокнами две передние трети его. Секреторная часть подходит к g?nglion submandibul?re и после перерыва в нем снабжает секреторными волокнами поднижнечелюстную и подъязычную слюнные железы.
После выхода из for?men stylomasto?deum от n. faci?lis отходят следующие мышечные ветви:
1. N. auricul?ris post?rior иннервирует m. auricul?ris post?rior и venter occipit?lis m. epicr?nii.
2. R?mus dig?stricus иннервирует заднее брюшко m. dig?stricus и m. stylohyo?deus.
3. Многочисленные ветви к мимической мускулатуре лица образуют и околоушной железе сплетение, pl?xus parot?deus. Ветви эти имеют в общем радиарное направление сзади наперед и, выходя из железы, идут на лицо и верхнюю часть шеи, широко анастомозируя с подкожными ветвями тройничного нерва. В них различают:
a) r?mi tempor?les к mm. auricul?res ant?rior et sup?rior, v?nter front?li m. epicr?mus и m. orbicul?ris ?culi;
б) r?mi zygom?tici к m. orbicul?ris ?culi и m. zygom?ticus;
в) r?mi bucc?les к мышцам в окружности рта и носа;
г) r?mus margin?lis mand?bulae — ветвь, идущую по краю нижней челюсти мышцам подбородка и нижней губы;
д) r?mus c?lli, которая спускается на шею и иннервирует m. plat?sma (рис. 329).
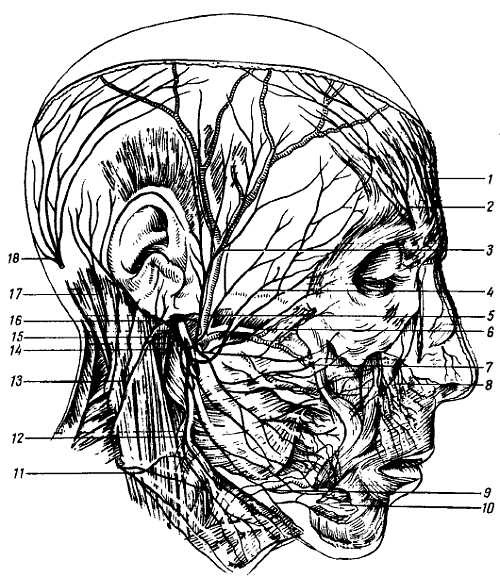
Рис. 329
1 — ветвь n. frontalis; 2 — n. supraorbitalis; 3 — ветви n. auriculotemporalis; 4 — r. zygomalicus n. facialis; 5 — n. auriculotemporaiis: 6, 7 — rr. buccales n. facialis; 8 — n. infraorbitalis; 9 — r. marginalis mandibulae; 10 — n. mentalis; 11 — n. transversus colli; 12 — r. colli n. facialis; 13 — n. auricularis magnus; 14 — r. communicans n. facialis к plexus cervicalis; 15 — n. facialis; 16 — n. auricularis posterior; 17 — n. occipitalis minor; 18 — n occipitalis major.
N. interm?dius, промежуточный нерв, является смешанным нервом. Он содержит афферентные (вкусовые) волокна, идущие к его чувствительному ядру (n?cleus solit?rius). и афферентные (секреторные, парасимпатические), исходящие из его вегетативною (секреторного) ядра (n?cleus salivat?rius sup?rior).
N. interm?dius выходит из мозга тонким стволиком между n. faci?lis и n. vestibulocochle?ris; пройдя некоторое расстояние между обоими этими нервами, он присоединяется к лицевому нерву, становится его составной частью, отчего n. interm?dius называют p?rtio interm?dia n. faci?lis. Далее он переходит в ch?rda t?mpani и n. petr?sus m?jor. Чувствительные его волокна возникают из отростков псевдоуниполярных клеток g?nglion gen?culi. Центральные отростки этих клеток идут в составе n. interm?dius в мозг, где оканчиваются в n?cleus solit?rius. Периферические отростки клеток проходят в ch?rda t?mpani, проводя вкусовую чувствительность от передней части языка и мягкого неба. Секреторные парасимпатические волокна от n. interm?dius начинаются в n?cleus salivat?rius sup?rior и направляются по ch?rda t?mpani к подъязычной и поднижнечелюстной железам (через посредство ganglion submandibul?re) и по n. petr?sus m?jor через g?nglion pterygopalat?num к железам слизистой оболочки носовой полости и неба. Слезная железа получает секреторные волокна из n. interm?dius через n. petr?sus m?jor, g?nglion pterygopalat?num и анастомоз второй ветви тройничного нерва с n. lacnmalis Таким образом можно сказать, что от n. lacrimalis (рис. 330).
Таким образом можно сказать, что от n. interm?dius иннервируются все железы, за исключением gl?ndula par?tis, получающей секреторные волокна от n. glossophar?ngeus.
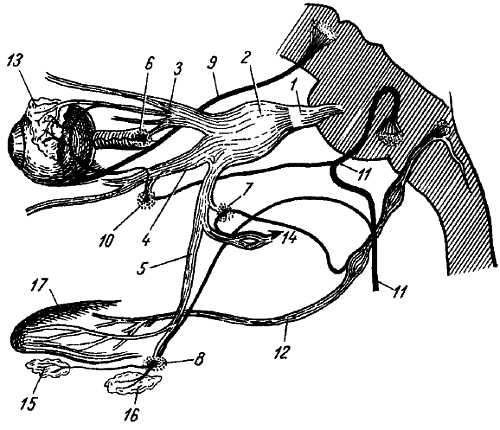
Рис. 330.
1 — n. trigeminus; 2 — gangl. trigeminale; 3 — n. ophthalmicus; 4 — n. maxillaris; 5 — n. mandibularis; 6 — gangl. ciliare; 7 — gangl. oticum; 8 — gangl. submandibulare; 9 — n. oculomotorius; 10 — gangl. pterygopalatinum; 11 — n. facialis; 12 — n. glossopharyngeus; 13 — gl. laerimalis; 14 — n. auriculotemporalis; 15 — gl. sublingualis; 16 — gl. submandibularis; 17 — lingua.
Преддверно-улитковый нерв (VIII)
N. vestibulocochle?ris, преддверно-улитковый, — обособившийся от лицевого нерва афферентный нерв, содержит соматически-чувствительные волокна, идущие от органа слуха и гравитации. Он состоит из двух частей — pars vestibul?ris и pars cochle?ris, которые по своим функциям различны, pars vestibul?ris является проводником импульсов от статического аппарата, заложенного в преддверии (vest?bulum) и полукружных протоках лабиринта внутреннего уха, a pars cochle?ris проводит слуховые импульсы от находящегося в улитке (c?chlea) спирального органа, воспринимающего звуковые раздражения.
Поскольку эти части чувствительные, каждая из них снабжена собственным нервным узлом, содержащим биполярные нервные клетки. Узел pars vestbul?ris, называемый g?nglion vestibul?re, лежит на дне внутреннего слухового прохода, а узел pars cochle?ris — g?nglion spir?le — помещается в улитке.
Периферические отростки биполярных клеток узлов оканчиваются в воспринимающих приборах вышеуказанных отделов лабиринта, о чем подробнее см.: «Орган слуха» и «Орган гравитации и равновесия». Центральные их отростки, выйдя из внутреннего уха через p?rus ac?sticus int?rnus направляются в составе соответствующих частей нерва к мозгу; они вступают в него сбоку лицевого нерва, достигая своих ядер; pars vestibul?ris — четырех и cochle?ris — двух ядер.
Языкоглоточный нерв (IX)
N. glossophar?ngeus, языкоглоточный нерв (рис. 331), нерв 3-й жаберной дуги, в процессе развития отделился от X пары нервов, n. v?gus.
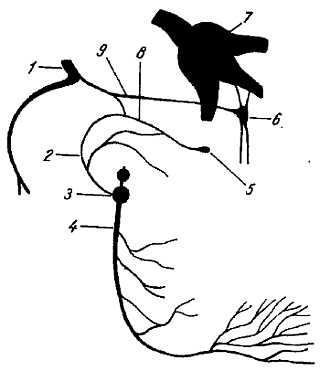
Рис. 331.
1 — n. facialis; 2 — n. tympanicus; 3 — gangl. inferius n. IX; 4 — n. glossopharyngeus; 5 — gangl. oticum; 6 — gangl. pterygopalatinum; 7 — gangl. trigeminale; 8 — n. petrosus minor; 9 — n. petrosus major.
Он содержит, в себе три рода волокон: 1) афферентные (чувствительные), идущие от рецепторов глотки, барабанной полости, слизистой оболочки языка (задней трети), миндалин и небных дужек; 2) эфферентные (двигательные), иннервирующие одну из мышц глотки (m. stylophar?ngeus); 3) эфферентные (секреторные), парасимпатические, для gl?ndula par?tis. Соответственно своим компонентам он имеет три ядра: n?cleus solit?rius, к которому приходят центральные отростки клеток 2 афферентных узлов — g?nglia sup?rius et inf?rius (см. ниже). Вегетативное (секреторное), парасимпатическое, ядро, n?cleus salivat?rius inf?rior (нижнее слюноотделительное ядро), состоит из клеток, рассеянных в form?tio reticul?ris около третьего ядра, двигательного, общего с n. v?gus n?cleus amb?guus (см. рис. 283). N. glossopharyng?us выходит своими корешками из продолговатого мозга позади оливы, над n. v?gus, и вместе с последним покидает череп через for?men jugul?re. В пределах последнего чувствительная часть нерва образует узел, g?nglion sup?rius, и по выходе из отверстия — другой узел, g?nglion inf?rius, лежащий на нижней поверхности пирамиды височной кости. Нерв спускается вниз, сначала между v. jugul?ris int?rna и a. car?tis int?rna, а затем огибает сзади m. stylophar?ngeus и по латеральной стороне этой мышцы подходит пологой дугой к корню языка, где он делится на свои конечные ветви (рис. 332).
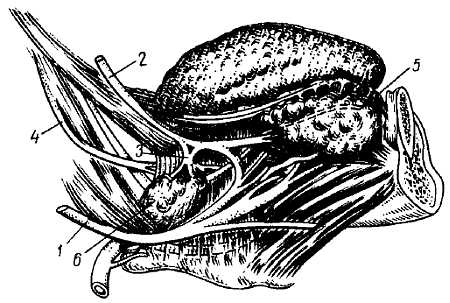
Рис. 332
1 — n. hypoglossus; 2 — n. lingualis; 3 — gangl. submandibulare; 4 — n. glossopharyngeus; 5 — gl. sublingualis; 6 — gl. submandibularis.
Ветви языкоглоточного нерва:
1. N. tymp?nicus отходит от g?nglion inf?rius и проникает в барабанную полость (c?vitas t?mpani), где образует сплетение, pl?xus tymp?nicus, к которому подходят ветви и от симпатического сплетения внутренней сонной артерии Это сплетение иннервирует слизистую оболочку барабанной полости и слуховой трубы. По выходе из барабанной полости через верхнюю стенку в виде n. petr?sus m?nor нерв проходит в одноименной бороздке, s?lcus n. petr?si min?ris, по передней поверхности пирамиды височной кости и достигает g?nglion ?ticum. Через этот нерв приносятся к g?nglion ?ticum исходящие из n?cleus salivat?rius inf?rior парасимпатические секреторные волокна для около-ушной железы. После перерыва в узле секреторные волокна подходят к железе в составе n. auriculotempo?alis от третьей ветви тройничного нерва.
2. Ramus m. stylophar?ngei к одноименной мышце.
3. R?mi tonsill?res к слизистой оболочке небных миндалин и дужек.
4. R?mi phar?ngei к глоточному сплетению (pl?xus phar?ngeus).
5. R?mi ligu?les (рис. 333), конечные ветви языкоглоточного нерва к слизистой оболочке задней трети языка, снабжающие ее чувствительными волокнами, среди которых проходят и вкусовые волокна к pap?llae vall?tae.
6. R. s?nus car?tici — чувствительный нерв к s?nus car?ticus (gl?mus car?ticum) (см. рис. 197).
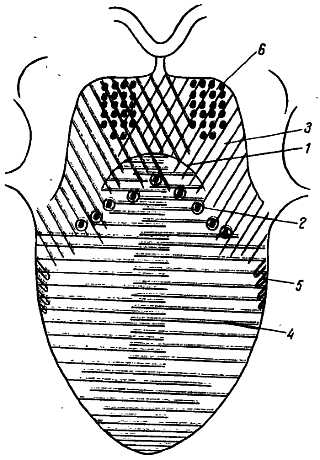
Рис. 333.
Впереди пограничной борозды (1) — желобовидные сосочки (2), сзади (3) — область иннервации n. glossopharyngeus; передняя часть языка (4) — область иннервации n. lingualis; по краям языка (5) — область иннервации n. glossopharyngeus; у корня языка (6) — область иннервации языка ветвью верхнего гортанного нерва (n. laryngeus superior).
Блуждающий нерв (X)
N. vagus, блуждающий нерв (см. рис. 272; рис. 334, 335), развившийся из 4-й и последующих жаберных дуг, называется так вследствие обширности его распространения.
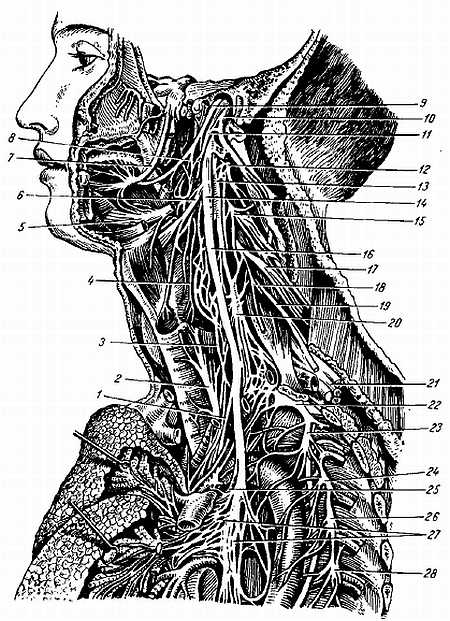
Рис. 334
1 — rr. cardiaci inferiores n. vagi; 2 — n. laryngeus inferior; 3 — rr. cardiaci superiores; 4 — plexus pharyngeus; 5 — n. hypoglossus; 6 — n. laryngeus superior; 7 — n. lingualis; 8 — rr. pharyngei n. vagi; 9 — n. glossopharyngeus; 10, 11 — ветви n. accessorii; 12, 15, 17, 19 — II, III, IV и V шейные спинномозговые нервы; 13 — gangl. cervicale superius; 14, 16 — n. vagus; 15 — n. phrenicus; 20 — gangl. cervicale medius; 21 — plexus brachialis; 22 — gangl. cervicale inferius; 23, 24, 26, 28 — II, III, IV и V грудные узлы truncus sympathicus; 25 — n. laryngeus recurrens; 27 — plexus pulmonalis.
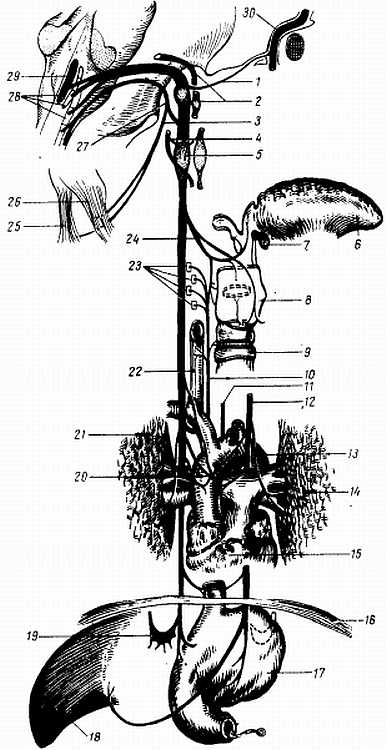
Рис. 335
1 — r. communicans n. vagi к n. facialis; 2 — n. glossopharyngeus; 3 — n. accessorius; 4 — r. communicans к n. hypoglossus; 5 — r. communicans к tr. sympathicus; 6 — lingua; 7 — os hyoideum; 8 — larynx; 9 — trachea; 10 — n. laryngeus recurrens dexter; 11 — n. laryngeus recurrens sinister; 12 — n. vagus sinister; 13 — aorta; 14 — pulmo sinister; 15 — cor; 16 — diaphragma; 17 — gaster; 18 — hepar; 19 — gangl. coeliacum dextrum; 20 — gangl. cardiacus; 21 — pulmo dexter; 22 — esophagus; 23 — разветвления n. laryngeus inferior в мышцах гортани; 24 — n. laryngeus superior; 25 — m. trapezius; 26 — m. sternocleidomastoideus; 27 — n. accessorius, проходящий через for. jugulare; 28 — ядра n. vagi и n. accessorii; 29 — ядро n. vagi; 30 — n. facialis.
Это самый длинный из черепных нервов. Своими ветвями блуждающий нерв снабжает дыхательные органы, значительную часть пищеварительного тракта (до c?lon sigmoideum), а также дает ветви к сердцу, которое получает от него волокна, замедляющие сердцебиение. N. v?gus содержит в себе троякого рода волокна:
1. Афферентные (чувствительные) волокна, идущие от рецепторов названных внутренностей и сосудов, а также от некоторой части твердой оболочки головного мозга и наружного слухового прохода с ушной раковиной к чувствительному ядру (n?cleus solit?rius).
2. Эфферентные (двигательные) волокна для произвольных мышц глотки, мягкого неба и гортани и исходящие от рецепторов этих мышц эфферентные (проприоцептивные) волокна. Эти мышцы получают волокна от двигательного ядра (n?cleus amb?guus).
3. Эфферентные (парасимпатические) волокна, исходящие из вегетативного ядра (n?cleus dors?lis n. v?gi). Они идут к миокарду сердца (замедляют сердцебиение) и мышечной оболочке сосудов (расширяют сосуды). Кроме того, в состав сердечных ветвей блуждающего нерва входит так называемый n. depr?ssor, который служит чувствительным нервом для самого сердца и начальной части аорты и заведует рефлекторным регулированием кровяного давления. Парасимпатические волокна иннервируют также трахею и легкие (суживают бронхи), пищевод, желудок и кишечник до c?lon sigmo?deum (усиливают перистальтику), заложенные в названных органах железы и железы брюшной полости — печень, поджелудочную железу (секреторные волокна), почки.
Парасимпатическая часть блуждающего нерва очень велика, вследствие чего он по преимуществу является вегетативным нервом, важным для жизненных функций организма. Блуждающий нерв представляет сложную систему, состоящую не только из нервных проводников разнородного происхождения, но и содержащую внутриствольные нервные узелки.
Волокна всех видов, связанные с тремя главными ядрами блуждающего нерва, выходят из продолговатого мозга в его s?lcus later?lis post?rior, ниже языкоглоточного нерва, 10–15 корешками, которые образуют толстый ствол нерва, покидающий вместе с языкоглоточным и добавочным нервами полость черепа через for?men jugul?re. В яремном отверстии чувствительная часть нерва образует небольшой узел — g?nglion sup?rius, а по выходе из отверстия — другое ганглиозное утолщение веретенообразной формы — g?nglion inf?rius. Тот и другой узел содержит псевдоуниполярные клетки, периферические отростки которых входят в состав чувствительных ветвей, идущих к названным узлам от рецепторов внутренностей и сосудов (g?nglion inf?rius) и наружного слухового прохода (g?nglion sup?rius), а центральные группируются в одиночный пучок, который заканчивается в чувствительном ядре, n?cleus solit?rius.
По выходе из полости черепа ствол блуждающего нерва спускается вниз на шею позади сосудов в желобке, сначала между v. jugul?ris int?rna и а. саr?tis int?rna, а ниже — между той же веной и a. c?rotis comm?nis, причем он лежит в одном влагалище с названными сосудами. Далее блуждающий нерв проникает через верхнюю апертуру грудной клетки в грудную полость, где правый его ствол располагается спереди a. subcl?via, а левый — на передней стороне дуги аорты. Спускаясь вниз, оба блуждающих нерва отходят сзади на той и другой сторонах корень легкого и сопровождают пищевод, образуя сплетения на его стенках, причем левый нерв проходит по передней стороне, а правый — по задней. Вместе с пищеводом оба блуждающих нерва проникают через hi?tus esoph?geus диафрагмы в брюшную полость, где образуют сплетения на стенках желудка. Стволы блуждающих нервов в утробном периоде располагаются симметрично по бокам пищевода. После поворота желудка слева направо левый v?gus, перемещается вперед, а правый назад, вследствие чего на передней поверхности разветвляется левый v?gus, а на задней — правый. От n. v?gus отходят следующие ветви:
А. В головной части (между началом нерва и g?nglion inf?rius):
1. R?mus men?ngeus к твердой оболочке головного мозга в области задней черепной ямки.
2. R?mus auricul?ris к задней стенке наружного слухового прохода и части кожи ушной раковины. Это единственная кожная веточка из черепных нервов, не относящаяся к n. trig?minus.
Б. В шейной части:
1. R?mi phar?ngei вместе с ветвями n. glossophar?ngeus и tr?ncus symp?thicus образуют сплетение, pl?xus phar?ngeus. Глоточные ветви блуждающего нерва иннервируют констрикторы глотки, мышцы небных дужек и мягкого неба (за исключением m. t?nsor v?li palat?ni). Глоточное сплетение дает еще чувствительные волокна к слизистой оболочке глотки.
2. N. lary?geus sup?rior снабжает чувствительными волокнами слизистую оболочку гортани выше голосовой щели, часть корня языка и надгортанника и двигательными — часть мышц гортани и нижний констриктор глотки.
3. R?mi card?aci cervic?les superi?res et inferi?res, частью могут выходить из n. lar?ngeus sup?rior, образуют сердечное сплетение.
В. В грудной части:
1. N. lar?ngeus rec?rrens, возвратный гортанный нерв, отходит в том месте, где n. v?gus лежит спереди дуги аорты (слева) или подключичной артерии (справа). На правой стороне этот нерв огибает снизу и сзади a. subcl?via, а на левой — также снизу и сзади дугу аорты и затем поднимается кверху в желобке между пищеводом и трахеей, давая им многочисленные ветви, r?mi esoph?gei и r?mi trache?les. Конец нерва, носящий название n. lar?ngeus inf?rior, иннервирует часть мышц гортани, слизистую оболочку ее ниже голосовых связок, участок слизистой оболочки корня языка около надгортанника, а также трахею, глотку и пищевод, щитовидную и вилочковую железы, лимфатические узлы шеи, сердце и средостение.
2. R?mi card?aci thor?cici берут начало от n. lar?ngeus rec?rrens и грудной части n. v?gus и идут к сердечному сплетению.
3. R?mi bronchi?les et trache?les вместе с ветвями симпатического ствола образуют на стенках бронхов сплетение, pl?xus pulmon?lis. За счет ветвей этого сплетения иннервируется мускулатура и железы трахеи и бронхов, а кроме того, оно содержит в себе и чувствительные волокна для трахеи, бронхов и легких.
4. R?mi esoph?gei идут к стенке пищевода.
Г. В брюшной части:
Сплетения блуждающих нервов, идущие по пищеводу, продолжаются на желудок, образуя выраженные стволы, tr?nci vag?les (передний и задний). Каждый tr?ncus vag?lis представляет собой комплекс нервных проводников не только парасимпатической, но также симпатической и афферентной анимальной нервной системы и содержит волокна обоих блуждающих нервов.
Продолжение левого блуждающего нерва, спускающегося с передней стороны пищевода на переднюю стенку желудка, образует сплетение, pl?xus g?stricus ant?rior, расположенное в основном вдоль малой кривизны, от которого отходят перемешивающиеся с симпатическими ветвями r?mi g?strici anteri?res к стенке желудка (к мышцам, железам и слизистой оболочке). Некоторые веточки через малый сальник направляются к печени. Правый n. v?gus на задней стенке желудка в области малой кривизны образует также сплетение, pl?xus g?stricus post?rior, дающее r?mi g?strici posteri?res; кроме того, б?льшая часть его волокон в виде r?mi coel?aci идет по тракту a. g?strica sin?stra к g?nglion coeli?cum, а отсюда по ветвям сосудов вместе с симпатическими сплетениями к печени, селезенке, поджелудочной железе, почкам, тонкой и толстой кишке до c?lon sigmo?deum. В случаях одностороннего или частичного повреждения X нерва нарушения касаются главным образом его анимальных функций. Расстройства висцеральной иннервации могут быть сравнительно нерезко выражены. Это объясняется, во-первых, тем, что в иннервации внутренностей имеются зоны перекрытия, а во-вторых, тем, что в стволе блуждающего нерва на периферии имеются нервные клетки — вегетативные нейроны, играющие роль в автоматической регуляции функций внутренностей.
Добавочный нерв (XI)
N. access?rius, добавочный нерв (см. рис. 272, 335), развивается из последних жаберных дуг, мышечный, содержит эфферентные (двигательные) и афферентные (проприоцептивные) волокна и имеет два двигательных ядра, заложенных в продолговатом и спинном мозге. Соответственно ядрам в нем различают церебральную и спинальную части. Церебральная часть выходит из продолговатого мозга тотчас ниже n. v?gus. Спинальная часть добавочного нерва формируется между передними и задними корешками спинномозговых нервов (С2—С5) и отчасти из передних корешков трех верхних шейных нервов, поднимается в виде нервного стволика вверх и присоединяется к церебральной части. Поскольку n. access?rius является отщепившейся частью блуждающего нерва, он и выходит с ним из полости черепа через for?men jugul?re иннервирует m. trap?zius и отделившийся от него m. sternocleidomasto?deus. Церебральная порция добавочного нерва в составе n. lar?ngeus rec?rrens идет для иннервации мышц гортани.
Спинальная порция добавочного нерва принимает участие в двигательной иннервации глотки, достигая ее мышц в составе блуждающего нерва, от которого добавочный нерв отщепился не полностью.
Общность и близость добавочного и языкоглоточного нервов с блуждающим объясняются тем, что IX, X и XI пары черепных нервов составляют одну группу жаберных нервов — группу вагуса, из которой выделился IX нерв и отщепился XI.
Нервы, развивающиеся в связи с головными миотомами
К этой группе относятся III, IV и VI пары черепных нервов, соответствующие передним корешкам спинномозговых нервов, выходящие из среднего мозга, в котором и заложены их ядра. Ядро VI пары вторично сместилось из среднего мозга в область ромбовидной ямки. Эти нервы являются двигательными корешками головных миотомов, поэтому они иннервируют мышцы глазного яблока, развившиеся из этих миотомов.
Глазодвигательный нерв (III)
N. oculomot?rius, глазодвигательный нерв, по развитию — двигательный корешок первого предушного миотома, является мышечным нервом. Содержит: 1) идущие из его соматически-двигательного ядра эфферентные (двигательные) волокна к большинству наружных мышц глазного яблока; 2) идущие от n?cleus access?rius парасимпатические волокна к внутренним глазным мышцам (m. sph?ncter pup?llae и m. cili?ris) (см. рис. 272). N. oculomot?rius выходит из мозга по медиальному краю ножки мозга, а затем идет до fiss?ra orbit?lis sup?rior, через которую входит в глазницу. Вступая в глазницу, делится на две ветви:
1. Верхнюю ветвь, r?mus sup?rior, к m. r?ctus sup?rior и m. lev?tor p?lpebrae superi?ris.
2. Нижнюю ветвь, r?mus inf?rior, к m. r?ctus inf?rior, m. r?ctus medi?lis и m. obl?quus inf?rior. От нижней ветви отходит к g?nglion cili?re корешок нерва, r?dix oculomot?ria, несущий парасимпатические волокна для m. sph?ncter pup?llae и m. cili?ris.
Блоковой нерв (IV)
N. trochle?ris, блоковой нерв, по развитию — двигательный корешок второго предушного миотома, является мышечным нервом и содержит идущие от его соматически-двигательного ядра эфферентные (двигательные) волокна к верхней косой мышце глаза. Выйдя с дорсальной стороны верхнего мозгового паруса, огибает латерально ножку мозга и через fiss?ra orbit?lis sup?rior входит в глазницу; оканчивается в m. obl?quus sup?rior (см. рис. 272).
Отводящий нерв (VI)
N. abducens, отводящий нерв, — двигательный корешок третьего предушного миотома, является мышечным нервом и содержит идущие из его соматически-двигательного ядра, заложенного в мосту, эфферентные (двигательные) волокна к латеральной прямой мышце глаза. Выходит из мозга у заднего края моста, проходит через fiss?ra orbit?lis sup?rior в глазницу и вступает в m. r?ctus later?lis (см. рис. 272).
Афферентные (пронриоцептивные) волокна для наружных глазных мышц, соответствующие эфферентным волокнам III, IV и VI нервов, идут в составе первой ветви V нерва, n. ophth?lmicus.
Многие авторы допускают наличие афферентных (проприоцептивных) волокон во всех трех двигательных нервах глазного яблока.
Нервы — производные мозга
К этой группе относятся nn. olfact?rii и n. ?pticus (см. рис. 272).
Обонятельные нервы (I)
Nn. olfact?rii, обонятельные нервы, развиваются из обонятельного мозга, возникшего в связи с рецептором обоняния. Они содержат висцерально-чувствительные волокна, идущие от органов восприятия химического раздражения. Поскольку нервы являются выростами переднего мозга, они не имеют узла, а представляют собой совокупность тонких нервных нитей, f?la olfact?ria, числом 15–20, которые являются центральными отростками обонятельных клеток, залегающих в r?gio olfact?ria слизистой оболочки носовой полости. F?la olfact?ria проходят через отверстия l?mina cribr?sa в верхней стенке носовой полости и затем оканчиваются в b?lbus olfact?rius, продолжающуюся в tr?ctus et trig?num olfact?rium.
Зрительный нерв (II)
N. ?pticus, зрительный нерв, в процессе эмбриогенеза вырастает как ножка глазного бокала из промежуточного мозга, а в процессе филогенеза связан со средним мозгом, возникающим в связи с рецептором света, чем и объясняются его прочные связи с этими отделами головного мозга. Он является проводником световых раздражений и содержит соматически-чувствительные волокна. Как производное мозга он не имеет узла, так же как и I пара черепных нервов, а входящие в его состав афферентные волокна составляют продолжение нейритов мультиполярных нервных клеток сетчатки глаза. Отойдя от заднего полюса глазного яблока, n. ?pticus покидает глазницу через can?lis ?pticus и, войдя в полость черепа вместе с таким же нервом другой стороны, образует перекрест, chi?sma ?pticum, лежащий в s?lcus chiasm?tis клиновидной кости (перекрест неполный, перекрещиваются лишь медиальные волокна нерва). Продолжением зрительного пути за хиазмой служит tr?ctus ?pticus, оканчивающийся в c?rpus genicul?tum later?le, p?lvinar th?lami и в верхнем холмике крыши среднего мозга (подробно см. «Орган зрения»). Между обеими сетчатками имеется связь посредством нервного пучка, идущего через передний угол перекреста. Эта связь аналогична комиссуральным связям полушарий мозга. Наличие указанной связи объясняет тот факт, что при повреждениях или заболеваниях одного глаза имеются выпадения поля зрения и в другом глазу.
Топография ядер черепных нервов, места входа их в мозг или выхода из него и из полости черепа представлены в табл. 2.
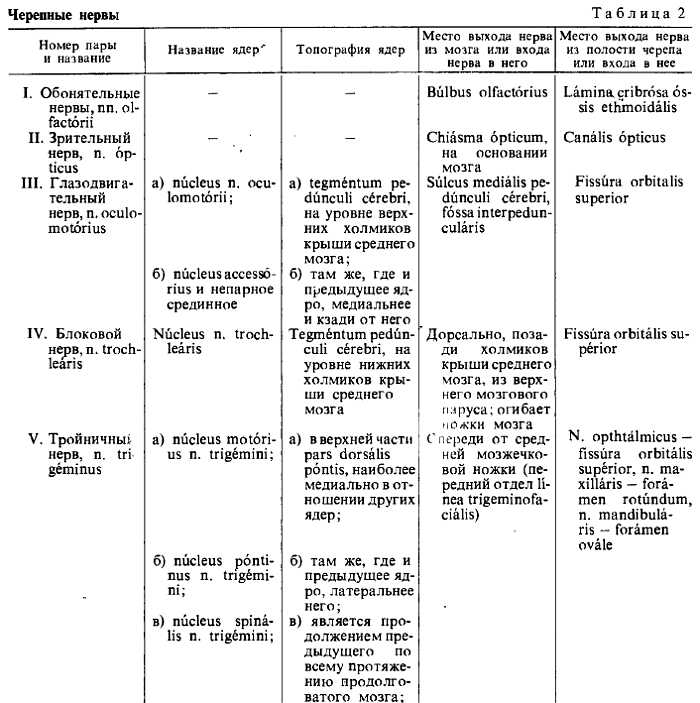

ПЕРИФЕРИЧЕСКАЯ ИННЕРВАЦИЯ СОМЫ
Каждый нерв распределяется посредством своих волокон в пределах определенной кожной или мышечной зоны, вследствие чего вся кожа и вся мускулатура могут быть поделены на зоны, соответствующие области разветвления данного кожного или мышечного нерва. Такая иннервация называется периферической, или зональной. Знание ее весьма важно для диагностики поражения нервов. На рис. 336 представлена периферическая иннервация кожи.
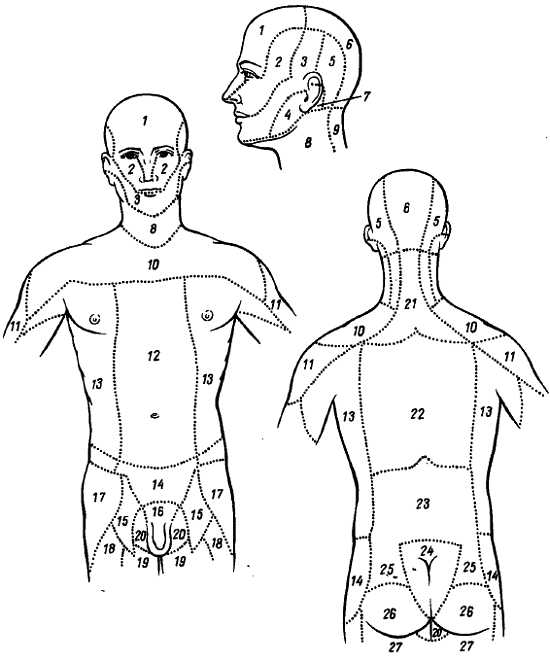
Рис. 336.
1 — n. ophthalmicus (V пара, 1-я ветвь); 2 — n. maxillaris (V пара, 2-я ветвь); 3 — n. mandibularis (V пара, 3-я ветвь); 4 — n. auricularis magnus (от plexus cervicalis); 5 — n. occipitalis minor (от plexus cervicalis); 6 — n. occipitalis major (C11); 7 — r. auricularis n. vagi; 8 — n. transversus coin (от plexus cervicalis); 9, 21 — задние ветви шейных нервов; 10 — rr. supraclaviculares mediales et intermedii (от plexus cervicalis); 11 — n. axillaris (от plexus brachialis); 12 — nn. intercostales (rr. cutanei anteriores); 13 — nn. intercostales (rr. cutanei laterales); 14 — n. iliohypogastricus (от plexus lumbalis); 15, 16 — r. femoralis и r. genitalis n. genitofemoralis (от plexus lumbalis); 17 — n. cutanens femoris lateralis (от plexus lumbalis); 18 — rr. cutanei anteriores (от n. femoralis); 19 — r. cutaneus n. obturatorii; 20 — rr. perineales n. cutanei femoris posterioris; 22 — rr. dorsales nn. thoracicorum; 23 — rr. dorsales nn. lumbalium; 24 — rr. dorsales nn. sacralium (от nn. clunium medii); 25 — nn. clunium siperiores; 26 — nn. clunium inferiores; 27 — n. cutaneus femoris posterior.
Что касается иннервации мышц, то о ней говорилось при описании каждой мышцы. Схема периферической иннервации мышц показана на рис. 337.
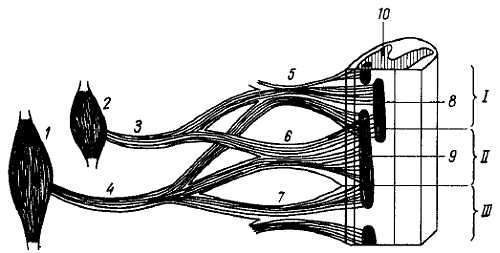
Рис. 337.
Мышца 2 иннервируется I и II спинно-мозговыми сегментами; мышца 1 иннервируется I, II и III спинномозговыми сегментами; 3, 4 — нервы; 5, 6, 7 — корешки; 8, 9 — группы клеток в передних рогах; 10 — задний рог.
Так как большинство нервов человеческого тела смешанные, то при их поражении расстройства чувствительности почти всегда сочетаются с двигательными. Область чувствительных расстройств в общем соответствует области, снабжаемой данным нервом. Однако соответствие оказывается далеко не полным, и зоны анестезии в действительности всегда значительно меньше указанных на схеме. Это зависит от частичного «перекрытия» данного нерва соседними и от их многочисленных связей. С этой точки зрения каждую область данного нерва можно разделить на три зоны:
1. Автономная зона, снабжаемая только данным нервом; при повреждении его наступает полная анестезия.
2. Смешанная зона, снабжаемая данным нервом и отчасти соседними; при повреждении данного нерва наблюдается частичное выпадение чувствительности — гипестезия.
3. Максимальная зона, снабжаемая полностью соседними нервами и лишь частично данным; при повреждении последнего чувствительность совсем не нарушается, так как сохраняется за счет соседних нервов.
Сегментарная, или корешковая, иннервация (рис. 338).
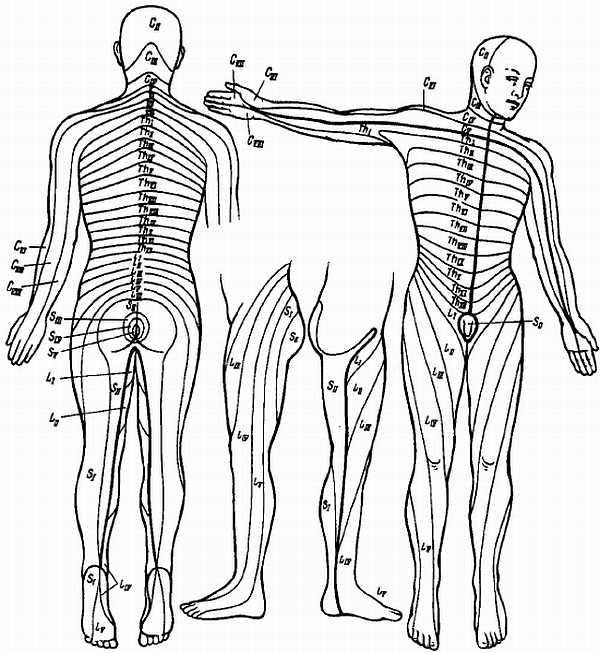
Рис. 338
Сообразно сегментарному строению организма каждый нервный сегмент (невромер) связан с соответствующим сегментом тела (сомитом). Поэтому каждый задний корешок спинномозгового нерва и каждый спинномозговой узел имеют отношение к иннервации того сегмента кожи (дерматома), который связан с ним в процессе эмбрионального развития. Точно так же и каждый передний корешок иннервирует те мышцы, которые произошли вместе с ним из данного сегмента (миотома) и вместе образуют нервно-мышечный сегмент. В результате вся кожа и вся мускулатура могут быть разделены на ряд последовательных корешковых зон, или поясов, иннервируемых соответствующими задними или передними нервными корешками. Это и составляет корешковую, или сегментарную, иннервацию тела, которая представлена на рис. 338. В отличие от зон периферической иннервации отдельных кожных нервов зоны корешковой иннервации имеют ту особенность, что волокна, относящиеся к одному заднему корешку или к одному сегменту, хотя бы они и шли в составе различных нервов, снабжают на коже определенную сплошную область, соответствующую всему данному нервному сегменту, или корешку, и потому называемую корешковым поясом. Корешковые, или сегментарные, зоны чувствительной иннервации идут на коже полосами, как показано на рис. 338. Поэтому в типичных случаях не представляет труда отличить сегментарное расстройство чувствительности от периферического. Так, при воспалении заднего корешка (радикулит) появляются опоясывающие боли или опоясывающий лишай, точно соответствующий данному корешковому поясу кожи. В практическом отношении важно знать, что соседние нервные сегменты целиком перекрывают друг друга, так что каждый сегмент кожи иннервируется тремя соседними нервными сегментами. Поэтому при перерезке одного корешка не удается обнаружить никаких расстройств чувствительности. Чтобы выпала чувствительность в одном сегменте кожи, надо перерезать три соседних нервных корешка, что следует учитывать при операциях. Также и при определении области поражения спинного мозга надо учитывать перекрытие сегментов и локализовать ее выше границ кожной анестезии на 1–2 сегмента.
ЗАКОНОМЕРНОСТИ РАСПРЕДЕЛЕНИЯ НЕРВОВ
1. Соответственно группировке органов тела вокруг нервной системы нервы расходятся в стороны от срединной линии, на которой располагается центральная нервная система (спинной и головной мозг).
2. Соответственно строению тела по принципу двусторонней симметрии нервы являются парными и идут симметрично.
3. Соответственно метамерному строению туловища нервы этой области сохраняют сегментарное строение (nn. intercost?les, ilioinguin?lis, iliohypog?stricus).
4. Нервы идут по кратчайшему расстоянию от места выхода из спинного или головного мозга к органу. Этим объясняется отхождение коротких ветвей к близлежащим органам и длинных — к отдаленным, идущих, однако, приблизительно по прямой линии, например n. ischi?dicus.
При перемещении органа от места первичной закладки в область окончательного расположения его после рождения нерв растет и следует за органом.
5. Нервы мышц отходят от сегментов спинного мозга, соответствующих миотомам, из которых происходит данная мышца. Поэтому даже при последующем перемещении мышцы она получает иннервацию от источника, расположенного вблизи первоначальной закладки. Этим объясняется иннервация трункопетальных мышц туловища, переместившихся на туловище с головы — от черепных нервов (n. access?rius), а с шеи — от шейного сплетения, или трункофугальных мышц конечностей от основного нервного сплетения данной конечности, например мышц пояса верхней конечности от плечевого сплетения. Этим же объясняется иннервация диафрагмы, закладывающейся на шее, от n. phr?nicus, происходящего из шейного сплетения.
Таким образом, по месту происхождения нерва можно определить область эмбрионального развития органа, ибо существует соответствие между происхождением нерва и местом закладки органов.
6. Если мышца представляет собой продукт слияния нескольких миотомов, то она иннервируется несколькими нервами (например, иннервация широких мышц живота межреберными нервами и ветвями поясничного сплетения). То же наблюдается в отношении висцеральных мышц, развивающихся из материала нескольких жаберных дуг. Так, переднее брюшко двубрюшной мышцы, возникающее из 1-й жаберной дуги, иннервируется тройничным нервом, а заднее брюшко, производное 2-й жаберной дуги, — лицевым нервом.
7. Поверхностные нервы (кожные) сопровождают подкожные вены, глубокие нервы сопровождают артерии, вены и лимфатические сосуды, образуя вместе с ними сосудисто-нервные пучки.
8. Нервы, заложенные в сосудисто-нервных пучках, как и эти последние, располагаются на сгибательных поверхностях данной области тела в защищенных, укрытых местах.
ВЕГЕТАТИВНАЯ (АВТОНОМНАЯ) НЕРВНАЯ СИСТЕМА
Выше отмечалась коренная качественная разница в строении, развитии и функции неисчерченных (гладких) и исчерченных (скелетных) мышц. Скелетная мускулатура участвует в реакции организма на внешние воздействия и отвечает на изменение среды быстрыми и целесообразными движениями. Гладкая мускулатура, заложенная во внутренностях и сосудах, работает медленно, но ритмично, обеспечивая течение жизненных процессов организма. Эти функциональные различия связаны с разницей в иннервации: скелетная мускулатура получает двигательные импульсы от анимальной, соматической части нервной системы, гладкая мускулатура — от вегетативной.
Вегетативная нервная система управляет деятельностью всех органов, участвующих в осуществлении растительных функций организма (питание, дыхание, выделение, размножение, циркуляция жидкостей), а также осуществляет трофическую иннервацию (И. П. Павлов).
Трофическая функция вегетативной нервной системы определяет питание тканей и органов применительно к выполняемой ими функции в тех или иных условиях внешней среды (адаптационно-трофическая функция).
Известно, что изменения в состоянии высшей нервной деятельности отражаются на функции внутренних органов и, наоборот, изменение внутренней среды организма оказывает влияние на функциональное состояние центральной нервной системы. Вегетативная нервная система усиливает или ослабляет функцию специфически работающих органов. Эта регуляция имеет тонический характер, поэтому вегетативная нервная система изменяет тонус органа. Так как одно и то же нервное волокно способно действовать лишь в одном направлении и не может одновременно повышать и понижать тонус, то сообразно с этим вегетативная нервная система распадается на два отдела, или части: симпатическую и парасимпатическую — pars symp?thica и pars parasymp?thica.
Симпатический отдел по своим основным функциям является трофическим. Он осуществляет усиление окислительных процессов, потребление питательных веществ, усиление дыхания, учащение деятельности сердца, увеличение поступления кислорода к мышцам.
Роль парасимпатического отдела охраняющая: сужение зрачка при сильном свете, торможение сердечной деятельности, опорожнение полостных органов.
Сравнивая область распространения симпатической и парасимпатической иннервации, можно, во-первых, обнаружить преобладающее значение одного какого-либо вегетативного отдела. Мочевой пузырь, например, получает в основном парасимпатическую иннервацию, и перерезка симпатических нервов не изменяет существенно его функции; только симпатическую иннервацию получают потовые железы, волосковые мышцы кожи, селезенка, надпочечники. Во-вторых, в органах с двойной вегетативной иннервацией наблюдается взаимодействие симпатических и парасимпатических нервов в форме определенного антагонизма. Так, раздражение симпатических нервов вызывает расширение зрачка, сужение сосудов, ускорение сердечных сокращений, торможение перистальтики кишечника; раздражение парасимпатических нервов приводит к сужению зрачка, расширению сосудов, замедлению сердцебиения, усилению перистальтики.
Однако так называемый антагонизм симпатической и парасимпатической частей не следует понимать статически, как противопоставление их функций. Эти части взаимодействующие, соотношение между ними динамически меняется на различных фазах функции того или иного органа; они могут действовать и антагонистически, и синергически.
Антагонизм и синергизм — две стороны единого процесса. Нормальные функции нашего организма обеспечиваются согласованным действием этих двух отделов вегетативной нервной системы. Эта согласованность и регуляция функций осуществляются корой головного мозга. В этой регуляции участвует и ретикулярная формация.
Автономия деятельности вегетативной нервной системы не является абсолютной и проявляется лишь в местных реакциях коротких рефлекторных дуг. Поэтому предложенный PNA термин «автономная нервная система» не является точным, чем и объясняется сохранение старого, более правильного и логичного термина «вегетативная нервная система». Деление вегетативной нервной системы на симпатический и парасимпатический отделы проводится главным образом на основании физиологических и фармакологических данных, но имеются и морфологические отличия, обусловленные строением и развитием этих отделов нервной системы.
Поэтому охарактеризуем сначала морфологические особенности вегетативной нервной системы в сравнении с анимальной. Опишем прежде всего центры вегетативной нервной системы (рис. 339).
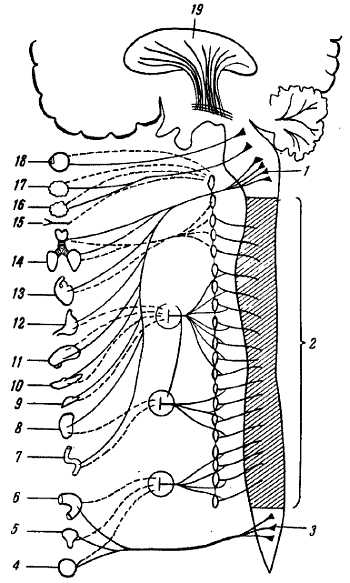
Рис. 339.
Пунктиром обозначены постганглионарные волокна симпатической системы, идущие к органам; непрерывной линией — преганглионарные волокна парасимпатической и симпатической систем. Область центров симпатической системы в спинном мозге заштрихована.
1 — центры парасимпатической системы в головном мозге (краниальный отдел); 2 — центры симпатической системы; 3 — центры парасимпатической системы в нижнем конце спинного мозга (крестцовый отдел); 4 — половые органы; 5 — мочевой пузырь; 6 — толстая кишка; 7 — тонкая кишка; 8 — почка; 9 — надпочечник; 10 — поджелудочная железа; 11 — печень; 12 — желудок; 13 — сердце; 14 — легкие; 15 — сосуды головы; 16, 17 — слюнные железы; 18 — глаз; 19 — полосатое тело.
Анимальные нервы выходят из мозгового ствола и спинного мозга на всем их протяжении сегментарно, причем эта сегментарность сохраняется частично и на периферии. Вегетативные нервы выходят только из нескольких отделов (очагов) центральной нервной системы. Имеются 4 таких очага, откуда выходят вегетативные нервы:
1. Мезэнцефалический отдел в среднем мозге (nucl. access?rius и непарное срединное ядро III пары черепных нервов).
2. Бульбарный отдел в продолговатом мозге и мосте (ядра VII, IX и X пар черепных нервов). Оба эти отдела объединяются под названием краниального.
3. Тораколюмбальный отдел в боковых рогах спинного мозга на протяжении сегментов СVIII, ThI—LIII.
4. Сакральный отдел в боковых рогах спинного мозга на протяжении сегментов SII—SIV.
Тораколюмбальный отдел относится к симпатической системе, а краниальный и сакральный — к парасимпатической (см. рис. 339; рис. 340).
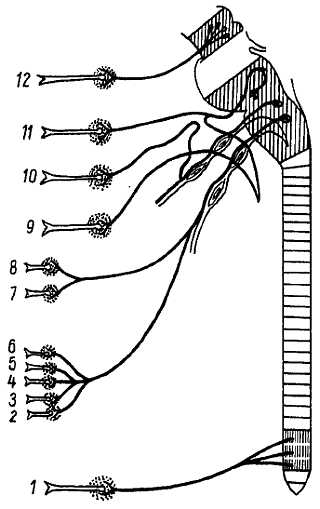
Рис. 340.
1 — подчревное сплетение: парасимпатические преганглионарные волокна подходят в составе тазовых нервов, постганглионарные направляются к прямой кишке, мочевому пузырю, половым органам. Постганглионарные волокна: 2 — к почкам; 3 — к поджелудочной железе; 4 — к печени; 5 — к кишечнику; 6 — к желудку; 7 — к бронхам; 8 — к сердцу (преганглионарные волокна 2–8 подходят в составе блуждающего нерва); 9 — поднижнечелюстной узел: преганглионарные волокна подходят по нерву, носящему название барабанной струны (ветвь лицевого нерва), постганглионарные направляются к поднижнечелюстной и подъязычной слюнным железам; 10 — ушной узел: преганлионарные волокна подходят по ветви языкоглоточного нерва; постганглионарные направляются к околоушной железе; 11 — крылонебный узел: преганглионарные волокна подходят по большому каменистому нерву (ветвь лицевого нерва), постганглионарные направляются к слезной железе и железам полости рта и носа; 12 — ресничный узел: преганглионарные волокна подходят по глазодвигательному нерву, постганглионарные направляются к сфинктеру зрачка и ресничной мышце глаза.
Над этими очагами доминируют высшие вегетативные центры, которые не являются симпатическими или парасимпатическими, а объединяют в себе регуляцию обоих отделов вегетативной нервной системы. К ним относится и ретикулярная формация. Они являются надсегментарными и расположены в стволе и плаще мозга, а именно:
1. Задний мозг: сосудодвигательный центр на дне IV желудочка; мозжечок, которому приписывают регуляцию ряда вегетативных функций (сосудодвигательные рефлексы, трофика кожи, скорость заживления ран и др.).
2. Средний мозг, серое вещество водопровода.
3. Промежуточный мозг: hypoth?lamus (t?ber ciner?um).
4. Конечный мозг: кора полушарий большого мозга.
Наибольшее значение для вегетативной регуляции имеет гипоталамическая область, которая является одним из самых древних отделов головного мозга, хотя и в ней различают более старые образования и филогенетически более молодые.
Гипоталамо-гипофизарная система, действуя с помощью инкретов гипофиза, является регулятором всех эндокринных желез.
Гипоталамическая область регулирует деятельность всех органов растительной жизни, объединяя и координируя их функции.
Объединение вегетативных и анимальных функций всего организма осуществляется в коре большого мозга, особенно в премоторной зоне.
Кора, будучи, по И. П. Павлову, комплексом корковых концов анализаторов, получает раздражения от всех органов, в том числе и от органов растительной жизни, и через посредство своих эфферентных систем, в том числе и вегетативной нервной системы, оказывает влияние на эти органы. Следовательно, существует двусторонняя связь коры и внутренностей — кортиковисцеральная связь. Благодаря этому все вегетативные функции подчиняются коре головного мозга, которая ведает всеми процессами организма.
Таким образом, вегетативная нервная система есть не автономная система, как это считали до И. П. Павлова, а специализированная часть единой нервной системы, подчиненная высшим отделам ее, включая и кору большого мозга. Поэтому, как и в анимальной нервной системе, в вегетативной можно различать центральный и периферический ее отделы. К центральному отделу относятся описанные выше очаги и центры в спинном и головном мозге, а к периферическому — нервные узлы, нервы, сплетения и периферические нервные окончания.
В последнее время установлено, что вегетативные узлы имеют свою афферентную иннервацию, благодаря которой они находятся под контролем центральной нервной системы.
Значительные отличия имеет рефлекторная дуга (рис. 341[рисунка нет]). Клеточное тело воспринимающего нейрона как для анимальной, так и для вегетативной нервной системы помещается в спинномозговом узле, g?nglion spin?le, куда стекаются афферентные пути как от органов животной жизни, так и от органов растительной жизни и который, таким образом, является смешанным анимально-вегетативным узлом. Клеточное тело вставочного нейрона вегетативной нервной системы в отличие от анимальной нервной системы помещается в боковых рогах спинного мозга. При этом аксон вставочного анимального нейрона, исходящий из клеток заднего рога, заканчивается в пределах спинного мозга среди клеток его передних рогов. Что же касается вставочного нейрона вегетативной нервной системы, то он в спинном мозге не заканчивается, а выходит за его пределы, к нервным узлам, расположенным на периферии. Выйдя из спинного мозга, аксон вставочного нейрона подходит или к узлам симпатического ствола, g?nglia tr?nci symp?thici, относящимся к симпатическому отделу вегетативной нервной системы (они образуют симпатический ствол), или волокна не заканчиваются в этих узлах, а направляются к предпозвоночным узлам, расположенным более периферично, между симпатическим стволом и органом (g?nglia coeli?ca, g?nglia mesent?rica). Эти узлы также относятся к симпатической системе. Наконец, волокна могут доходить, не прерываясь, до узлов, лежащих или около органа (околоорганные узлы, например g?nglia cili?re, ?ticum и др.), или в толще органа (внутриорганные, интрамуральные узлы); и те и другие называют конечными узлами (g?nliga termin?lia). Они относятся к парасимпатическому отделу вегетативной нервной системы. Кроме макроскопически видимых обособленных узлов, по ходу вегетативных нервов встречаются мигрировавшие сюда в ходе эмбрионального развития небольшие группы эффекторных нейронов — микроганглии. Все волокна, идущие до узлов первого, второго или третьего порядка и являющиеся аксонами промежуточного нейрона, называются предузловыми волокнами, r?mi preganglion?res. Они покрыты миелином.
Третий, эффекторный, нейрон анимальной рефлекторной дуги помещается в передних рогах спинного мозга, а эффекторный нейрон вегетативной рефлекторной дуги вынесен в процессе развития из центральной нервной системы в периферическую, ближе к рабочему органу, и располагается в вегетативных нервных узлах. Из такого расположения эффекторных нейронов на периферии вытекает главный признак вегетативной нервной системы — двухнейронность эфферентного периферического пути: первый нейрон — вставочный; тело его лежит в вегетативных ядрах черепных нервов или боковых рогах спинного мозга, а нейрит идет к узлу; второй — эфферентный, тело которого лежит в узле, а нейрит достигает рабочего органа. Эффекторные нейроны симпатических нервов начинаются в g?nglia tr?nci symp?thici (узлы первого порядка) или g?nglia interm?dia (узлы второго порядка), а для парасимпатических нервов — в около- или внутриорганных узлах, g?nglia termin?lia (третьего порядка); так как в названных узлах осуществляется связь вставочных и эфферентных нейронов, то отмеченная разница между симпатическим и парасимпатическим отделами вегетативной нервной системы связана именно с этими нейронами.
Аксоны эфферентных вегетативных нейронов почти лишены миелина — безмиелиновые (серые). Они составляют послеузловые волокна, r?mi postganglion?res. Послеузловые волокна симпатической нервной системы, отходящие от узлов симпатического ствола, расходятся в двух направлениях. Одни волокна идут к внутренностям и составляют висцеральную часть симпатической системы. Другие волокна образуют r?mi communic?ntes grisei, соединяющие симпатический ствол с анимальными нервами. В составе последних волокна достигают соматических органов (аппарата движения и кожи), в которых иннервируют непроизвольную мускулатуру сосудов и кожи, а также железы.
Совокупность описанных эфферентных вегетативных волокон, идущих от узлов симпатического ствола до органов сомы, составляет соматическую часть симпатического отдела. Такая структура обеспечивает функцию вегетативной нервной системы, которая регулирует обмен веществ всех частей организма применительно к непрерывно изменяющимся условиям среды и условиям функционирования (работы) тех или иных органов и тканей.
Соответственно этой наиболее универсальной своей функции, связанной не с какими-либо отдельными органами и системами, а со всеми частями, со всеми органами и тканями организма, вегетативная нервная система и морфологически характеризуется универсальным, повсеместным распространением в организме.
Следовательно, симпатический отдел иннервирует не только внутренности, но и сому, обеспечивая в ней обменные и трофические процессы.
В результате каждый орган, по И. П. Павлову, находится под тройным нервным контролем, в связи с чем он различает три вида нервов: 1) функциональные, осуществляющие функцию данного органа; 2) сосудодвигательные, обеспечивающий доставку крови к органу, и 3) трофические, регулирующие усвоение из доставленной крови питательных веществ.
Висцеральная часть симпатического отдела содержит все эти три вида нервов для внутренностей, а соматическая часть — только сосудодвигательные и трофические. Что же касается функциональных нервов для органов сомы (скелетная мускулатура и др.), то они идут в составе соматической, анимальной, нервной системы.
Таким образом, основное отличие эфферентной части вегетативной нервной системы от эфферентной части анимальной заключается в том, что анимальные, соматические, нервные волокна, выйдя из центральной нервной системы, идут до рабочего органа, нигде не прерываясь, тогда как вегетативные волокна на своем пути от мозга до рабочего органа прерываются в одном из узлов первого, второго или третьего порядка. Вследствие этого эфферентный путь вегетативной нервной системы разбивается на две части, из которых он и состоит: предузловые миелиновые волокна, rami preganglionares, и послеузловые, лишенные миелина (безмиелиновые) волокна, r?mi postganglion?res.
Наличие узлов в эфферентной части рефлекторной дуги составляет характерный признак вегетативной нервной системы, отличающий ее от анимальной (см. рис. 341).
Определенные отличия имеют и нервы. Афферентные пути вегетативной нервной системы не имеют характера макроскопически видимых нервов, а их волокна идут в составе других нервов (nn. spl?nchnici m?jor et minor, n. v?gus, задние корешки и др.). При этом для симпатического отдела характерно то, что связанная с ним чувствительная иннервация может распространяться на значительные расстояния и, следовательно, симпатический отдел может рассматриваться как система окольной иннервации.
Так, например, афферентные спинальные нервные волокна, участвующие в формировании чревного сплетения, иннервирующего органы брюшной полости, происходят из многочисленных спинномозговых узлов (СV—LIII). Это обстоятельство определяет множественность и многосегментарность путей и источников афферентной иннервации органов брюшной полости. Этим же объясняется и то, что чувство боли от внутренностей может передаваться как по вегетативным, так и по анимальным нервам.
Существуют также собственные афферентные нейроны вегетативной нервной системы, замыкающиеся в вегетативных ганглиях, которые могут рассматриваться как периферические центры.
Наряду с этим имеется не только диффузность в отношении распределения чувствительных нейронов и их волокон, но и преимущественное участие определенных спинномозговых узлов в иннервации внутренностей. Следовательно, среди источников и путей афферентной иннервации внутренностей можно выделить основные и дополнительные. Это деление тесно связано с представлением об окольных путях афферентной спинальной иннервации внутренностей. Окольные пути в патологических условиях (перерыв спинного мозга и др.) могут играть роль компенсаторных путей, возмещающих функцию нарушенных основных путей, компенсаторных приспособлений в виде «перекрытия» в афферентной иннервации органов.
Что же касается эфферентных путей вегетативной нервной системы, то они образуют ясно выраженные нервы и узлы. Поэтому можно говорить о двух центробежных путях единой нервной системы: один путь — это анимальные, соматические, двигательные нервы, а другой — вегетативные. Вегетативные нервы образуют сплетения вокруг кровеносных сосудов, вместе с которыми они подходят и входят в органы. Наличие сплетений вокруг сосудов составляет характерный признак вегетативной нервной системы, отличающий ее от анимальной.
Как уже отмечалось, вегетативная нервная система характеризуется универсальным, повсеместным распространением в организме. Она имеет широкую область эфферентной иннервации, охватывающую все органы и ткани тела, не исключая и скелетной мускулатуры (последнюю она тонизирует). В этом и состоит морфологическая особенность вегетативной нервной системы в противоположность анимальной, которая иннервирует центробежными волокнами только скелетные мышцы, т. е. имеет сравнительно ограниченную область эфферентной иннервации.
Для понимания строения необходимо учитывать развитие вегетативной нервной системы.
Гладкая мускулатура беспозвоночных регулируется ганглиозно-сетевидной нервной системой, которая, кроме этой специальной функции, регулирует также и обмен веществ. Приспособление уровня обмена веществ к изменяющейся функции органов называется адаптацией (adapt?re — прилаживать), а соответственная функция нервной системы — адаптационно-трофической (Л. А. Орбели). Адаптационно-трофическая функция есть наиболее общая и весьма древняя функция нервной системы, существовавшая у примитивных предков позвоночных. В дальнейшем ходе эволюции сильнее всего прогрессировали аппарат движения (развитие твердого скелета и скелетной мускулатуры) и органы чувств, т. е. органы животной жизни. Поэтому та часть нервной системы, которая была связана с ними, т. е. анимальная часть нервной системы, претерпела наиболее резкие изменения и приобрела новые признаки, в частности: изоляция волокон при помощи миелиновых оболочек, большая скорость проведения возбуждения (100–120 м/с). Напротив, органы растительной жизни претерпели более медленную и менее прогрессивную эволюцию, поэтому связанная с ними часть нервной системы сохранила за собой наиболее общую функцию — адаптационно-трофическую. Эта часть нервной системы и есть вегетативная нервная система.
Наряду с некоторой специализацией она сохранила ряд древних примитивных черт: отсутствие у большинства нервных волокон миелиновых оболочек (безмиелиновые волокна), меньшая скорость проведения возбуждения (0,3-10 м/с), а также меньшая концентрация и централизация эффекторных нейронов, оставшихся разбросанными на периферии, в составе ганглиев, нервов и сплетений. При этом эффекторный нейрон оказался расположенным вблизи рабочего органа или даже в толще его.
Такое периферическое расположение эффекторного нейрона обусловило главную морфологическую особенность вегетативной нервной системы — двухнейронность эфферентного периферического пути, состоящего из вставочного и эффекторного нейронов.
С появлением туловищного мозга (у бесчерепных) возникающие в нем импульсы адаптации идут по вставочным нейронам, обладающим большей скоростью возбуждения; выполняется же адаптация непроизвольной мускулатурой и железами, к которым подходят эффекторные нейроны, отличающиеся медленной проводимостью. Это противоречие разрешается в процессе эволюции благодаря развитию специальных нервных узлов, в которых устанавливаются контакты вставочных нейронов с эффекторными, причем один вставочный нейрон вступает в связь со многими эффекторными (примерно 1:32). Этим достигается переключение импульсов с миелиновых волокон, обладающих большой скоростью проведения раздражений, на безмиелиновые, обладающие малой скоростью. В результате весь эфферентный периферический путь вегетативной нервной системы разбивается на две части — предузловую и послеузловую, а сами узлы становятся трансформаторами темпов возбуждения с быстрых на медленные.
У низших рыб, когда образуется головной мозг, в нем развиваются центры, объединяющие деятельность органов, вырабатывающих внутреннюю среду организма.
Так как в этой деятельности, кроме гладкой мускулатуры, принимает участие и скелетная (исчерченная), то возникает потребность в координации работы гладких и поперечно-полосатых мышц. Например, жаберные крышки приводятся в движение скелетной мускулатурой, так же и у человека в акте дыхания участвует как гладкая мускулатура бронхов, так и скелетные мышцы грудной клетки. Такую координацию осуществляет развивающийся в заднем мозге специальный рефлекторный аппарат в виде системы блуждающего нерва (бульбарный отдел парасимпатической части вегетативной нервной системы).
В центральной нервной системе возникают и другие образования, которые подобно блуждающему нерву выполняют функцию координации совместной деятельности скелетной мускулатуры, обладающей быстрой скоростью возбуждения, и гладкой мускулатуры и желез, обладающих медленной скоростью. Сюда относится та часть глазодвигательного нерва, которая осуществляет при помощи исчерченных и неисчерченных мышц глаза стандартную установку ширины зрачка, аккомодации и конвергенции соответственно силе освещения и расстоянию до рассматриваемого объекта по тем же принципам, как это делает фотограф (мезэнцефалический отдел парасимпатической части вегетативной нервной системы). Сюда относится и та часть крестцовых нервов (II–IV), которые осуществляют стандартную функцию тазовых органов (мочевого пузыря и прямой кишки) — опорожнение, в которой участвуют как непроизвольные мышцы этих органов, так и произвольные мышцы таза и брюшного пресса — сакральный отдел парасимпатической части вегетативной нервной системы.
В среднем и промежуточном мозге развился центральный адаптационный аппарат в виде серого вещества вокруг водопровода и серого бугра (hypoth?lamus).
Наконец, в коре мозга возникли центры, объединяющие высшие анимальные и вегетативные функции.
Развитие вегетативной нервной системы в онтогенезе (эмбриогенезе) идет иначе, чем в филогенезе.
Вегетативная нервная система возникает из общего с анимальной частью источника — нейроэктодермы, чем доказывается единство всей нервной системы.
Из общего зачатка нервной системы выселяются симпатобласты, которые скапливаются в определенных местах, образуя сначала узлы симпатического ствола, а затем промежуточные узлы, а также нервные сплетения. Отростки клеток симпатического ствола, объединяясь в пучки, образуют r?mi communic?ntes gr?sei.
Сходным образом развивается и часть вегетативной нервной системы в области головы. Зачатки парасимпатических узлов выселяются из продолговатого мозга или ганглиозной пластинки и совершают дальнюю миграцию вдоль ветвей тройничного, блуждающего и других нервов, оседая по их ходу или образуя интрамуральные ганглии.
СИМПАТИЧЕСКАЯ ЧАСТЬ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Исторически симпатическая часть возникает как сегментарный отдел, поэтому и у человека она частично сохраняет сегментарный характер строения.
ЦЕНТРАЛЬНЫЙ ОТДЕЛ СИМПАТИЧЕСКОЙ ЧАСТИ
Центральный отдел симпатической части располагается в боковых рогах спинного мозга на уровне СVIII, ThI—LIII, в subst?ntia interm?dia later?lis. От него отходят волокна, иннервирующие непроизвольные мышцы внутренних органов, органов чувств (глаза), железы. Кроме того, здесь располагаются сосудодвигательные и потоотделительные центры. Считают (и это подтверждается клиническим опытом), что различные отделы спинного мозга оказывают влияние на трофику, терморегуляцию и обмен веществ.
ПЕРИФЕРИЧЕСКИЙ ОТДЕЛ СИМПАТИЧЕСКОЙ ЧАСТИ
Периферический отдел симпатической части образуется прежде всего двумя симметричными стволами, tr?nci symp?thici d?xter et sinister, расположенными по бокам позвоночника на всем его протяжении от основания черепа до копчика, где оба ствола своими каудальными концами сходятся в одном общем узле. Каждый из этих двух симпатических стволов слагается из ряда нервных узлов первого порядка, соединяющихся между собой посредством продольныхмежузловых ветвей, r?mi intergangliori?res, состоящих из нервных волокон. Кроме узлов симпатических стволов (g?nglia tr?nci symp?thici), в состав симпатической системы входят указанные выше g?nglia interm?dia.
Симпатический ствол, начиная с верхнего шейного узла, содержит также элементы парасимпатической части вегетативной и даже анимальной нервных систем.
Отростки клеток, заложенных в боковых рогах тораколюмбального отдела спинного мозга, выходят из спинного мозга через передние корешки и, отделившись от них, идут в составе r?mi communic?ntes ?lbi к симпатическому стволу (см. рис. 341). Здесь они или соединяются синапсом с клетками узлов симпатического ствола, или же, пройдя через его узлы без перерыва, достигают одного из промежуточных узлов. Это так называемый преганглионарный путь. От узлов симпатического ствола или (если там не было перерыва) от промежуточных узлов отходят безмиелиновые волокна постганглионарного пути, направляющиеся к кровеносным сосудам и внутренностям.
Поскольку симпатическая часть имеет соматическую часть, она связана со спинномозговыми нервами, обеспечивающими иннервацию сомы. Эта связь осуществляется посредством серых соединительных ветвей, r?mi communic?ntes gr?sei, которые представляют собой участок постганглионарных волокон на протяжении от узлов симпатического ствола до n. spin?lis. В составе r?mi communic?ntes gr?sei и спинномозговых нервов постганглионарные волокна распространяются в сосудах, железах и мышцах, поднимающих волосы кожи туловища и конечностей, а также в скелетной мускулатуре, обеспечивая ее трофику и тонус.
Таким образом, симпатическая часть соединяется с анимальной нервной системой посредством двоякого рода соединительных ветвей: белых и серых, r?mi communic?ntes ?lbi et gr?sei. Белые соединительные ветви (миелиновые) имеют в своем составе преганглионарные волокна. Они идут от центров симпатической части через передние корешки к узлам симпатического ствола. Поскольку центры лежат на уровне грудных и верхних поясничных сегментов, то и r?mi communic?ntes ?lbi имеются лишь в пределах от I грудного до III поясничного спинномозгового нерва. R?mi communic?ntes gr?sei, постганглионарные волокна, обеспечивают вазомоторные и трофические процессы сомы; они соединяют симпатический ствол со спинномозговыми нервами на всем его протяжении. Шейный отдел симпатического ствола имеет связь и с черепными нервами. Следовательно, все сплетения анимальной нервной системы содержат в составе своих пучков и нервных стволов волокна симпатической части, чем подчеркивается единство этих систем.
Симпатический ствол
Каждый из двух симпатических стволов подразделяют на четыре отдела: шейный, грудной, поясничный (или брюшной) и крестцовый (или тазовый).
Шейный отдел простирается от основания черепа до шейки I ребра; располагается позади сонных артерий на глубоких мышцах шеи. В его состав входят три шейных симпатических узла: верхний, средний и нижний.
G?nglion cervic?le sup?rius является самым крупным узлом симпатического ствола, имея длину около 20 мм и ширину 4–6 мм. Лежит он на уровне II и части III шейных позвонков позади внутренней сонной артерии и медиально от n. v?gus.
G?nglion cervic?le m?dium небольшой величины, располагается обыкновенно в месте перекреста a. thyro?dea inf?rior с сонной, артерией, нередко отсутствует или может распадаться на два узелка.
G?nglion cervic?le inf?rius довольно значительной величины, расположен позади начальной части позвоночной артерии; нередко сливается с I, а иногда и II грудным узлом, образуя общий шейно-грудной, или звездчатый, узел, g?nglion cervicothor?cicum s. g?nglion stell?tum.
От шейных узлов отходят нервы для головы, шеи и груди. Их можно разделить на восходящую группу, направляющуюся к голове, на нисходящую — опускающуюся к сердцу, и группу для органов шеи.
Нервы для головы отходят от верхнего и нижнего шейных узлов и делятся на группу, проникающую в полость черепа, и группу, подходящую к голове снаружи.
Первая группа представлена n. car?ticus int?rnus, отходящим от верхнего шейного узла, и n. vertebr?lis, отходящим от нижнего шейного узла. Оба нерва, сопровождая одноименные артерии, образуют вокруг них сплетения: pl?xus car?ticus int?rnus и pl?xus vertebr?lis; вместе с артериями они проникают в полость черепа, где анастомозируют между собой и дают ветви к сосудам мозга, оболочкам, гипофизу, стволам III, IV, V, VI пар черепных нервов и барабанному нерву.
PI?xus car?ticus int?rnus продолжается в pl?xus cavern?sus, которое окружает а. car?tis int?rna на участке прохождения ее через s?nus cavern?sus.
Ветви сплетений распространяются, кроме самой внутренней сонной артерии, также по ее разветвлениям. Из ветвей pl?xus car?ticus int?rnus следует отметить n. petr?sus prof?ndus, который присоединяется к n. petr?sus m?jor и вместе с ним образует n. can?lis pterygo?dei, подходящий через одноименный канал к g?nglion pterygopalat?num.
Вторая группа симпатических нервов головы, наружная, составляется двумя ветвями верхнего шейного узла, nn. car?tici ext?rni, которые, образовав сплетение вокруг наружной сонной артерии, сопровождают ее разветвления на голове. От этого сплетения отходит стволик к ушному узлу, gangl. oticum; от сплетения, сопровождающего лицевую артерию, отходит ветвь к поднижнечелюстному узлу, gangl. submandibulare.
Через посредство ветвей, входящих в сплетения вокруг сонной артерии и ее ветвей, верхний шейный узел дает волокна к сосудам (вазоконстрикторы) и железам головы: потовым, слезной, слизистым и слюнным, а также к мышцам волос кожи и к мышце, расширяющей зрачок (см. «Орган зрения»), m. dilat?tor pup?llae. Центр расширения зрачка, c?ntrum ciliospin?le, находится в спинном мозге на уровне от VIII шейного до II грудного сегмента.
Органы шеи получают нервы от всех трех шейных узлов; кроме того, часть нервов отходит от межузловых участков шейного отдела симпатического ствола, а часть — от сплетений сонных артерий.
Веточки от сплетений следуют по ходу ветвей наружной сонной артерии, носят одноименные названия и вместе с ними подходят к органам, в силу чего число отдельных симпатических сплетений равно числу артериальных ветвей. Из нервов, отходящих от шейной части симпатического ствола отмечают гортанно-глоточные ветви от верхнего шейного узла — r?mi laryngophar?ngei, которые частью идут с n. lar?ngeus sup?rior (ветвь n. v?gi) к гортани, частью спускаются к боковой стенке глотки; здесь они вместе с ветвями языкоглоточного, блуждающего и верхнего гортанного нервов образуют глоточное сплетение, pl?xus phar?ngeus.
Нисходящая группа ветвей шейной части симпатического ствола представлена nn. card?aci cervic?les sup?rior, m?dius et inf?rior, отходящими от соответствующих шейных узлов. Шейные сердечные нервы спускаются в грудную полость, где вместе с симпатическими грудными сердечными нервами и ветвями блуждающего нерва участвуют в образовании сердечных сплетений (см. иннервацию сердца).
Грудной отдел симпатического ствола располагается впереди шеек ребер, прикрыт спереди плеврой. В его состав входят 10–12 узлов более или менее треугольной формы. Грудной отдел характеризуется присутствием белых соединительных ветвей, r?mi communic?ntes ?lbi, соединяющих передние корешки спинномозговых нервов с узлами симпатического ствола. Ветви грудного отдела: 1) nn. card?aci thor?cici отходят от верхних грудных узлов и участвуют в образовании pl?xus card?acus (подробное описание сердечных сплетений см. при описании сердца); 2) r?mi communic?ntes gr?sei, безмиелиновые — к межреберным нервам (соматическая часть симпатического отдела); 3) r?mi pulmon?les — к легким, образуют pl?xus pulmon?lis; 4) r?mi a?rtici образуют сплетение на грудной аорте, pl?xus a?rticus thor?cicus, и частью на пищеводе, pl?xus esoph?geus, а также на грудном протоке (во всех указанных сплетениях принимает участие и n. v?gus); 5) nn. spl?nchnici m?jor et m?nor, большой и малый внутренностные нервы; n. spl?nchnicus m?jor начинается несколькими корешками, отходящими от V–IX грудных узлов; корешки n. spl?nchnicus m?jor идут в медиальном направлении и сливаются на уровне IX грудного позвонка в один общий ствол, проникающий через промежуток между мышечными пучками ножек диафрагмы в брюшную полость, где он входит в состав pl?xus coel?acus; n. spl?nchnicus minor начинается от X–XI грудных узлов и также входит в pl?xus coel?acus, проникая через диафрагму с большим внутренностным нервом. В этих нервах проходят сосудосуживающие волокна, как это видно из того обстоятельства, что при перерезке этих нервов сосуды кишечника сильно переполняются кровью; в nn. spl?nchnici содержатся волокна, тормозящие движение желудка и кишок, а также волокна, служащие проводниками ощущений от внутренностей (афферентные волокна симпатической части).
Поясничный, или брюшной, отдел симпатического ствола состоит из четырех, иногда из трех узлов. Симпатические стволы в поясничном отделе расположены на более близком расстоянии один от другого, чем в грудной полости, так что узлы лежат на переднебоковой поверхности поясничных позвонков вдоль медиального края m. ps?as m?jor. R?mi communic?ntes ?lbi имеются только с двумя или тремя верхними поясничными нервами.
От брюшного отдела симпатического ствола на всем протяжении отходит большое количество ветвей, которые вместе с nn. spl?nchnici m?jor et m?nor и брюшными отделами блуждающих нервов образуют самое большое непарное чревное сплетение, pl?xus coel?acus. В формировании чревного сплетения участвуют также многочисленные спинномозговые узлы (С5—L3), аксоны их нейроцитов (Д. М. Голуб, 1963). Оно лежит на передней полуокружности брюшной аорты, позади поджелудочной железы, и окружает начальные части чревного ствола (tr?ncus c?eliacus) и верхней брыжеечной артерии. Сплетение занимает участок между почечными артериями, надпочечниками и аортальным отверстием диафрагмы и включает парный чревный узел, g?nglion c?eliacum, и иногда непарный верхний брыжеечный узел, g?nglion mesent?ricum sup?rius.
От чревного сплетения отходит ряд меньших парных сплетений к диафрагме, надпочечникам, почкам, а также pl?xus testicul?ris (ov?ricus), следующих по ходу одноименных артерий. Имеется также ряд непарных сплетений к отдельным органам по стенкам артерий, название которых они носят. Из последних верхнее брыжеечное сплетение, pl?xus mesent?ricus sup?rior, иннервирует поджелудочную железу, тонкую и толстую кишку до половины протяжения поперечной ободочной.
Вторым главным источником иннервации органов полости живота является сплетение на аорте, pl?xus a?rticus abdomin?lis. составленное из двух стволов, отходящих от чревного сплетения, и веточек от поясничных узлов симпатического ствола. От аортального сплетения отходит нижнее брыжеечное сплетение, pl?xus mesent?ricus inf?rior, для поперечной и нисходящей части ободочной кишки, сигмовидной и верхних отделов r?ctum (pl?xus r?ctaIs superi?r). У места отхождения pl?xus mesent?ricus inf?rior располагается одноименный узел, gangl. mesent?ricum inf?rius. Его постганглионарные волокна идут в тазе в составе nn. hypog?strici.
Аортальное сплетение продолжается вначале в непарное верхнее подчревное сплетение, pl?xus hypog?stricus sup?rior, которое у мыса раздваивается и переходит в сплетение таза, или нижнее подчревное сплетение (pl?xus hypog?stricus inf?rior s. pl?xus pelv?nus).
Волокна, происходящие из верхних поясничных сегментов, по своей функции являются сосудодвигательными (вазоконстрикторами) для полового члена, двигательными для матки и сфинктера мочевого пузыря.
Крестцовый, или тазовый, отдел имеет обычно четыре узла; располагаясь на передней поверхности крестца вдоль медиального края передних крестцовых отверстий, оба ствола книзу постепенно сближаются друг с другом и затем оканчиваются в одном общем непарном узле — g?nglion ?mpar; находящемся на передней поверхности копчика. Узлы тазового отдела, как и поясничного, связаны между собой не только продольными, но и поперечными стволиками.
От узлов крестцового отдела симпатического ствола отходит ряд ветвей, которые соединяются с ветвями, отделяющимися от нижнего брыжеечного сплетения, и образуют пластинку, протягивающуюся от крестца к мочевому пузырю; это так называемое нижнее подчревное, или тазовое, сплетение, рl?хus hypog?stricus inf?rior s. pl?xus pelv?nus. Сплетение имеет свои узелки — g?nglia pelv?na. В сплетении различают несколько отделов: 1) передненижний отдел, в котором выделяют верхнюю часть, иннервирующую мочевой пузырь. — pl?xus vesic?lis, и нижнюю, снабжающую у мужчин предстательную железу (pl?xus pr?staticus), семенные пузырьки и семявыносящий проток (pl?xus deferenti?lis) и пещеристые тела (nn. cavern?si p?nis); 2) задний отдел сплетения снабжает прямую кишку (pl?xus rect?les m?dii et inferi?res). У женщин выделяют еще средний отдел, нижняя часть которого дает ветви к матке и влагалищу (pl?xus uterovagin?lis), пещеристым телам клитора (nn. cavern?si clit?ridis), а верхняя — к матке и яичникам.
От узлов крестцового отдела симпатического ствола отходят соединительные ветви, r?mi communic?ntes, присоединяющиеся к спинномозговым нервам, иннервирующим нижнюю конечность. Эти соединительные ветви составляют соматическую часть симпатического отдела вегетативной нервной системы, иннервирующую нижнюю конечность. В составе r?mi communic?ntes и спинномозговых нервов нижней конечности находятся постганглионарные волокна, которые распространяются в сосудах, железах и мышцах волос кожи, а также в скелетной мускулатуре, обеспечивая ее трофику и тонус.
ПАРАСИМПАТИЧЕСКАЯ ЧАСТЬ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Парасимпатическая часть (см. рис. 340) исторически развивается как надсегментарный отдел, и поэтому центры ее располагаются не только в спинном мозге, но и в головном.
Центры парасимпатической части
Центральная часть парасимпатического отдела состоит из головного, или краниального, отдела и спинномозгового, или сакрального, отдела. Некоторые авторы считают, что парасимпатические центры располагаются в спинном мозге не только в области крестцовых сегментов, но и в других отделах его, в частности в пояснично-грудном отделе между передним и задним рогом, в так называемой интермедиарной зоне. Центры дают начало эфферентным волокнам передних корешков, вызывающих расширение сосудов, задержку потоотделения и торможение сокращения непроизвольных мышц волос в области-туловища и конечностей.
Краниальный отдел в свою очередь состоит из центров, заложенных в среднем мозге (мезэнцефалическая часть), и в ромбовидном мозге — в мосту и продолговатом мозге (бульбарная часть).
1. Мезэнцефалическая часть представлена n?cleus access?rius n. oculomot?rii и срединным непарным ядром, за счет которых иннервируется мускулатура глаза — m. sphincter pupillae и m. cili?ris.
2. Бульбарная часть представлена nucleus salivat?rius sup?rior n. faci?lis (точнее, n. interm?dius), n?cleus salivat?rius inf?rior n. glossophar?ngei и n?cleus dors?lis n. v?gi (см. соответствующие нервы).
Сакральный отдел (рис. 342). Парасимпатические центры лежат в спинном мозге, в subst?ntia intermedialater?lis бокового рога на уровне II–IV крестцовых сегментов.
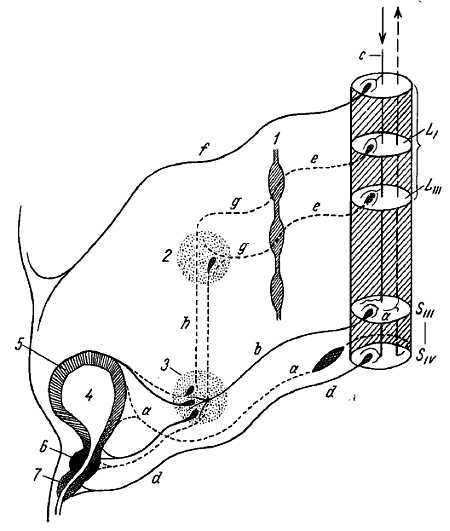
Рис. 342.
а — чувствительный путь к спинному мозгу; б — n. splanchnicus pelvinus; с — кортико-спинальный путь; d — n. pudendus; е — rr. communicantes; f — n. iliohypogastricus; g — nn. mesenterici; h — nn. hypogastrici; 1 — симпатический ствол; 2 — gangl. mesentericum inferius; 3 — plexus hypogastricus и расположенные поблизости терминальные узлы парасимпатической системы; 4 — vesica urinaria; 5 — m. detrusor vesicae; 6 — m. sphincter vesicae; 7 — m. sphincter urethrae.
Периферический отдел парасимпатической части
Периферическая часть краниального отдела парасимпатической системы представлена: 1) преганглионарными волокнами, идущими в составе III, VII, IX и X пар черепных нервов (возможно, и в составе I и XI); 2) терминальными узлами, расположенными вблизи органов, а именно: g?nglia cili?re, pterygopalat?num, submandibul?re, ?ticum, и 3) постганглионарными волокнами; постганглионарные волокна имеют или самостоятельный ход, как, например, nn. cili?res br?ves, отходящие от g?nglion cili?re, или идут в составе каких-либо нервов, как, например, постганглионарные волокна, отходящие от g?nglion ?ticum и идущие в составе n. auriculotempor?lis. Некоторые авторы указывают, что парасимпатические волокна выходят также и из других сегментов спинного мозга и идут через передние корешки, направляясь к стенкам туловища и конечностей.
Периферическая часть сакрального отдела парасимпатической системы представлена волокнами, которые в составе передних корешков II–IV крестцовых нервов и далее в составе их передних ветвей, образующих pl?xus sacr?lis (анимальное сплетение), входят в малый таз. Здесь они отделяются от сплетения и в виде nn. spl?nchnici pelv?ni направляются к pl?xus hypog?stricus inf?rior, иннервируя вместе с последним тазовые внутренности: прямую кишку с c?lon sigmoideum, мочевой пузырь, наружные и внутренние половые органы. Раздражение nn. spl?nchnici pelv?ni вызывает сокращение прямой кишки и мочевого пузыря (m. detr?sor v?sicae) с ослаблением их сфинктеров. Волокна симпатического подчревного сплетения задерживают опорожнение этих органов; они же возбуждают сокращение матки, тогда как nn. spl?nchnici pelv?ni его тормозят. Nn. spl?nchnici pelv?ni содержат в себе еще сосудорасширяющие волокна (nn. erig?ntes) для c?rpora cavern?sa p?nis et clit?ridis, обусловливающие эрекцию. Парасимпатические волокна, отходящие от сакрального отдела спинного мозга, идут в тазовые сплетения не только в составе nn. erig?ntes и nn. spl?nchnici pelv?ni, но и в составе n?rvus pud?ndus (преганглионарные волокна). Половой нерв является сложным нервом, содержащим в своем составе, кроме анимальных волокон, также и вегетативные (симпатические и парасимпатические), входящие в нижнее подчревное сплетение. Симпатические волокна, отходящие от узлов крестцового отдела симпатического ствола в качестве постганглионарных, присоединяются к половому нерву в полости малого таза и проходят через нижнее подчревное сплетение к тазовым органам.
К парасимпатической нервной системе относится также так называемая интрамуральная нервная система.
В стенках ряда полостных органов находятся нервные сплетения, содержащие мелкие узлы (терминальные) с ганглиозными клетками и безмиелиновыми волокнами, — ганглиозно-сетевидная, или интрамуральная, система.
Интрамуральная система особенно выражена в пищеварительном тракте, где она представлена несколькими сплетениями.
1. Мышечно-кишечное сплетение, pl?xus myent?ricus — между продольной и кольцевой мускулатурой пищеварительной трубки.
2. Подслизистое сплетение, pl?xus submuc?sus, находящееся в подслизистой основе. Последнее переходит в сплетение желез и ворсинок.
К периферии от названных сплетений располагается диффузная нервная сеть. К сплетениям подходят нервные волокна от симпатической и парасимпатической систем. В интрамуральных сплетениях происходит переключение предузловых волокон парасимпатической системы на послеузловые.
Интрамуральные сплетения, как и экстраорганные сплетения полостей туловища, являются по своему составу смешанными. В последнее время в интрамуральных сплетениях пищеварительного тракта обнаружены и клетки симпатической природы.
КРАТКИЙ ОБЗОР ВЕГЕТАТИВНОЙ ИННЕРВАЦИИ ОРГАНОВ[37]
Иннервация глаза. В ответ на определенные зрительные раздражения, идущие от сетчатки, осуществляются конвергенция и аккомодация зрительного аппарата.
Конвергенция глаз — сведение зрительных осей обоих глаз на рассматриваемом предмете — происходит рефлекторно, сочетанным сокращением поперечно-полосатых мышц глазного яблока. Этот рефлекс, необходимый для бинокулярного зрения, связан с аккомодацией глаза. Аккомодация — способность глаза ясно видеть предметы, находящиеся от него на различных расстояниях, зависит от сокращения мышц глаза — m. cili?ris и m. sph?ncter pup?llae. Поскольку деятельность мускулатуры глаза осуществляется совместно с сокращением его поперечно-полосатых мышц, вегетативная иннервация глаза будет рассмотрена вместе с анимальной иннервацией его двигательного аппарата.
Афферентным путем от мышц глазного яблока (проприоцептивная чувствительность) являются, по данным одних авторов, сами анимальные нервы, иннервирующие данные мышцы (III, IV, VI черепные нервы), по данным других — n. ophth?lmicus (n. trig?mini).
Центры иннервации мышц глазного яблока — ядра III, IV, и VI пар. Эфферентный путь — III, IV и VI черепные нервы. Конвергенция глаза осуществляется, как указывалось, сочетанным сокращением мышц обоих глаз.
Надо иметь в виду, что изолированных движений одного глазного яблока вообще не существует. В любых произвольных и рефлекторных движениях всегда участвуют оба глаза. Эта возможность сочетанного движения глазных яблок (взора) обеспечивается особой системой волокон, связывающей между собой ядра III, IV и VI нервов и носящей название медиального продольного пучка.
Медиальный продольный пучок начинается от ядра в ножках мозга, соединяется с ядрами III, IV, VI нервов при помощи коллатералей и направляется по мозговому стволу вниз в спинной мозг, где заканчивается, по-видимому, в клетках передних рогов верхних шейных сегментов. Благодаря этому движения глаз сочетаются с движениями головы и шеи.
Иннервация гладких мышц глаза — m. sph?ncter pup?llae и m. cili?ris происходит за счет парасимпатической системы, иннервация m. dilat?tor pup?llae — за счет симпатической. Афферентными путями вегетативной системы являются n. oculomot?rius и n. ophth?lmicus.
Эфферентная парасимпатическая иннервация. Преганглионарные волокна идут из добавочного ядра глазодвигательного нерва (мезэнцефалический отдел парасимпатической нервной системы) в составе n. oculomot?rius и по его r?dix oculomot?ria достигают g?nglion cili?re (рис. 343), где и оканчиваются. В ресничном узле начинаются постганглионарные волокна, которые через nn. cili?res br?ves доходят до ресничной мышцы и сфинктера зрачка. Функция: сужение зрачка и аккомодация глаза к дальнему и близкому видению.
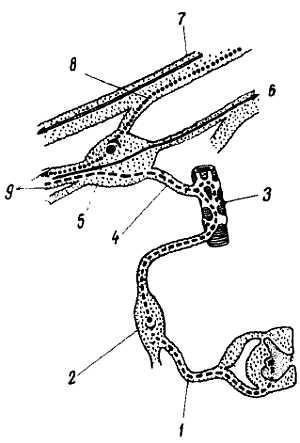
Рис. 343
1 — r. commuriicans albus; 2 — gangl. cervicale superius; 3 — a. ophthalmica; 4 — r. sympathicus от gangl. ciliare; 5 — gangl. ciliare; 6 — n. nasociliaris; 7 — n. oculomotorius; 8 — radix oculomotoria (парасимпатические преганглионарные волокна); 9 — nn. ciliares breves.
Эфферентная симпатическая иннервация. Преганглионарные волокна идут из клеток subst?ntia intermediolater?lis боковых рогов последнего шейного и двух верхних грудных сегментов (СVIII—ThII c?ntrum ciliospin?le), выходят через две верхние грудные r?mi communic?ntes ?lbi, проходят в составе шейного отдела симпатического ствола и оканчиваются в верхнем шейном узле. Постганглионарные волокна идут в составе n. car?ticus int?rnus в полость черепа и вступают в pl?xus car?ticus int?rnus и pl?xus opht?lmicus, после этого часть волокон проникает в r?mus comm?nicans, соединяющуюся с n. nasocili?ris, и n?rvi cili?res l?ngi, а часть направляется к ресничному узлу, через который проходит, не прерываясь. в n?rvi cili?res br?ves. И те и другие симпатические волокна, проходящие через длинные и короткие ресничные нервы, направляются к дилататору зрачка. Функция: расширение зрачка, а также сужение сосудов глаза.
Иннервация желез — слезной и слюнных. Афферентным путем для слезной железы является n. lacrim?lis (ветвь n. ophth?lmicus от n. trig?mini), для поднижнечелюстной и подъязычной — n. lingu?lis (ветвь n. mandibul?ris от n. trig?mini) и ch?rda t?mpani (ветвь n. interm?dius), для околоушной — n. auriculotempor?lis и n. glossophar?ngeus.
Эфферентная парасимпатическая иннервация слезной железы. Центр лежит в верхнем отделе продолговатого мозга и связан с ядром промежуточного нерва (n?cleus salivat?rius sup?rior). Преганглионарные волокна идут в составе n. interm?dius. далее n. petr?sus major до g?nglion pterygopalat?num (рис. 344). Отсюда начинаются постганглионарные волокна, которые в составе n. maxill?ris и далее его ветви, n. zygom?ticus, через связи с n. lacrim?lis достигают слезной железы.
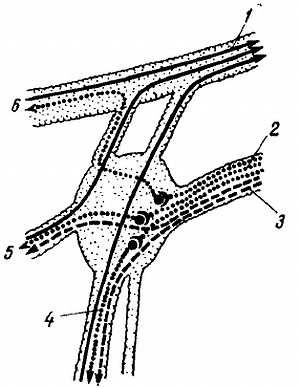
Рис. 344
1 — n. maxillaris; 2 — n. petrosus major; 3 — n. petrosus profundus; 4 — nn. palatini; 5 — nn. nasales posteriores; 6 — n. zygomaticus.
Эфферентная парасимпатическая иннервация поднижиечелюстной и подъязычной желез. Преганглионарные волокна идут от n?cleus salivat?rius sup?rior в составе n. interm?dius, далее ch?rda t?mpani и n. lingu?lis до g?nglion submandibul?re, откуда начинаются постанглионарные волокна, достигающие желез.
Эфферентная парасимпатическая иннервация околоушной железы. Преганглионарные волокна идут от n?cleus salivat?rius inf?rior в составе n. glossophar?ngeus, далее n. tymp?nicus, n. petr?sus m?nor до g?nglion ?ticum (рис. 345). Отсюда начинаются постганглионарные волокна, идущие к железе в составе n. auriculotempor?lis. Функция: усиление секреции слезной и названных слюнных желез; расширение сосудов желез.
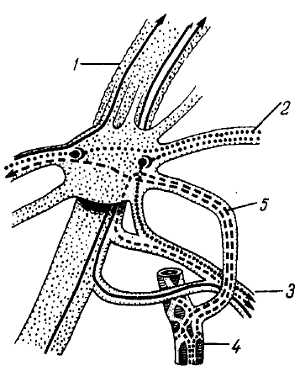
Рис. 345.
1 — n. mandibularis; 2 — n. petrosus minor; 3 — n. auriculotemporalis; 4 — a. meningea media; 5 — симпатические волокна.
Эфферентная симпатическая иннервация всех названных желез. Преанглионарные волокна начинаются в боковых рогах верхних грудных сегментов спинного мозга и заканчиваются в верхнем шейном узле симпатического ствола. Постганглионарные волокна начинаются в названном узле и доходят до слезной железы в составе pl?xus car?ticus int?rnus, до околоушной — в составе pl?xus car?ticus ext?rnus и до поднижнечелюстной и подъязычной желез — через pl?xus car?ticus ext?rnus и затем через pl?xus faci?lis. Функция: задержка отделения слюны (сухость во рту); слезотечение (влияние не резкое).
Иннервация сердца (рис. 346). Афферентные пути от сердца идут в составе n. v?gus, а также в среднем и нижнем шейных и грудных сердечных симпатических нервах. При этом по симпатическим нервам проводится чувство боли, а по парасимпатическим — все остальные афферентные импульсы.
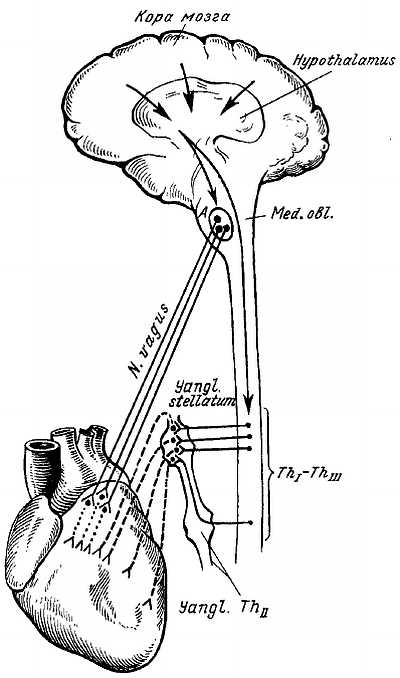
Рис. 346.
А — ядро сердечных волокон блуждающего нерва в продолговатом мозге: ТhI-ТhIII — сегменты спинного мозга, в которых расположены клетки, отдающие симпатические нервные волокна к сердцу. Сплошные линии — преганглионарные волокна блуждающего и симпатических нервов: пунктирные — постгаглионарные волокна блуждающего нерва в сердце; прерывистые линии — постганглионарные симпатические волокна, идущие к сердцу. Стрелками показано направление влияния коры мозга, передаваемое через гипоталамус на ядра блуждающего нерва и симпатические центры
Эфферентная парасимпатическая иннервация. Пpeанглионарные волокна начинаются в дорсальном вегетативном ядре блуждающего нерва и идут в составе последнего, его сердечных ветвей (r?mi cardi?ci n. v?gi) и сердечных сплетений (см. иннервацию сердца) до внутренних узлов сердца, а также узлов околосердечных полей. Постганглионарные волокна исходят от этих узлов к мышце сердца. Функция: торможение и угнетение деятельности сердца; сужение венечных артерий.
Эфферентная симпатическая иннервация. Преганглионарные волокна начинаются из боковых рогов спинного мозга 4–5 верхних грудных сегментов, выходят в составе соответственных r?mi communic?ntes ?lbi и проходят через симпатический ствол до пяти верхних грудных и трех шейных узлов. В этих узлах начинаются постганглионарные волокна, которые в составе сердечных нервов, nn. cardi?ci cervic?les sup?rior, m?dius et inf?rior и nn. cardi?ci thor?cici, достигают сердечной мышцы. Перерыв осуществляется только в g?nglion stell?tum. Сердечные нервы содержат в своем составе преганглионарные волокна, которые переключаются на постганглионарные в клетках сердечного сплетения. Функция: усиление работы сердца (это установил И. П. Павлов в 1888 г., назвав симпатический нерв усиливающим) и ускорение ритма (это впервые установил И. Ф. Цион в 1866 г.), расширение венечных сосудов.
Иннервация легких и бронхов. Афферентными путями от висцеральной плевры являются легочные ветви грудного отдела симпатического ствола, от париетальной плевры — nn. intercost?les и n. phr?nicus, от бронхов — n. v?gus.
Эфферентная парасимпатическая иннервация. Преганглионарные волокна начинаются в дорсальном вегетативном ядре блуждающего нерва и идут в составе последнего и его легочных ветвей к узлам pl?xus pulmon?lis, а также к узлам, расположенным по ходу трахеи, бронхов и внутри легких. Постганглионарные волокна направляются от этих узлов к мускулатуре и железам бронхиального дерева. Функция: сужение просвета бронхов и бронхиол и выделение слизи.
Эфферентная симпатическая иннервация. Преганглионарные волокна выходят из боковых рогов спинного мозга верхних грудных сегментов (ThII—ThVI) и проходят через соответствующие r?mi communic?ntes ?lbi и симпатический ствол к звездчатому и верхним грудным узлам. От последних начинаются постганглионарные волокна, которые проходят в составе легочного сплетения к бронхиальной мускулатуре и кровеносным сосудам. Функция: расширение просвета бронхов; сужение.
Иннервация желудочно-кишечного тракта (до сигмовидной кишки), поджелудочной железы, печени. Афферентные пути от указанных органов идут в составе n. v?gus, n. spl?nchnicus m?jor et minor, pl?xus hep?ticus, pl?xus coel?acus, грудных и поясничных спинномозговых нервов и в составе n. phr?nicus.
По симпатическим нервам передается чувство боли от этих органов, по n. v?gus — другие афферентные импульсы, а от желудка — чувство тошноты и голода.
Эфферентная парасимпатическая иннервация. Преганглионарные волокна из дорсального вегетативного, ядра блуждающего нерва проходят в составе последнего до терминальных узлов, находящихся в толще названных органов. В кишечнике это клетки кишечных сплетений (pl?xus myent?ricus, submuc?sus). Постганглионарные волокна идут от этих узлов к гладким мышцам и железам. Функция: усиление перистальтики желудка, расслабление сфинктера привратника, усиление перистальтики кишок и желчного пузыря, расширение сосудов. В составе блуждающего нерва имеются волокна, возбуждающие и тормозящие секрецию.
Эфферентная симпатическая иннервация. Преганглионарные волокна выходят из боковых рогов спинного мозга V–XII грудных сегментов, идут по соответствующим r?mi communic?ntes ?lbi в симпатический ствол и далее без перерыва в составе nn. spl?nchnici maj?res (VI–IX) до промежуточных узлов, участвующих в образовании чревного, верхнего и нижнего брыжеечных сплетений (g?nglia coeliaca и g?nglion mesent?ricum sup?rius et inf?rius). Отсюда возникают постганглионарные волокна, идущие в составе pl?xus coeliacus и pl?xus mesent?ricus sup?rior к печени, p?ncreas, к тонкой кишке и к толстой до середины c?lon transv?rsum; левая половина c?lon transv?rsum и c?lon desc?ndens иннервируется из pl?xus mesent?ricus inf?rior. Указанные сплетения снабжают мускулатуру и железы названных органов. Функция: замедление перистальтики желудка, кишок и желчного пузыря, сужение просвета кровеносных сосудов и угнетение секреции желез.
К этому нужно заметить, что задержка движений желудка и кишечника достигается также и тем, что симпатические нервы вызывают активное сокращение сфинктеров: sph?ncter pyl?ri, сфинктеры кишечника и др.
Иннервация сигмовидной и прямой кишки и мочевого пузыря. Афферентные пути идут в составе pl?xus mesent?ricus inf?rior, pl?xus hypog?stricus sup?rior et inf?rior и в составе nn. spl?nchnici pelv?ni.
Эфферентная парасимпатическая иннервация. Преганглионарные волокна начинаются в боковых рогах спинного мозга II–IV крестцовых сегментов и выходят в составе соответствующих передних корешков спинномозговых нервов. Далее они идут в виде nn. spl?nchnici pelv?ni до внутриорганных узлов названных отделов толстой кишки и околоорганных узлов мочевого пузыря. В этих узлах начинаются постганглионарные волокна, которые достигают гладкой мускулатуры названных органов. Функция: возбуждение перистальтики сигмовидной и прямой кишки, расслабление m. sph?ncter ?ni int?rnus, сокращение m. detr?sor v?sicae и расслабление m. sph?ncter v?sicae.
Эфферентная симпатическая иннервация. Преганглионарные волокна идут от боковых рогов поясничного отдела спинного мозга через соответствующие передние корешки в r?mi communic?ntes ?lbi, проходят, не прерываясь, через симпатический ствол и достигают g?nglion mesent?ricum inf?rius. Здесь начинаются постганглионарные волокна, идущие в составе nn. hypog?strici до гладкой мускулатуры названных органов. Функция: задержка перистальтики сигмовидной и прямой кишки и сокращение внутреннего сфинктера прямой кишки. В мочевом пузыре симпатические нервы вызывают расслабление m. detr?sor v?sicae и сокращение сфинктера мочевого пузыря.
Иннервация половых органов: симпатическая — см. «Симпатический ствол», парасимпатическая — см. «Периферический отдел парасимпатической системы». Иннервация других внутренних органов приводится после их описания.
Иннервация кровеносных сосудов. Степень иннервации артерий, капилляров и вен неодинакова. Артерии, у которых более развиты мышечные элементы в t?nica m?dia, получают более обильную иннервацию, вены — менее обильную; v. c?va inf?rior и v. p?rtae занимают промежуточное положение.
Более крупные сосуды, расположенные внутри полостей тела, получают иннервацию от ветвей симпатического ствола, ближайших сплетений вегетативной нервной системы и прилежащих спинномозговых нервов; периферические же сосуды стенок полостей и сосуды конечностей получают иннервацию от проходящих поблизости нервов. Нервы, подходящие к сосудам, идут сегментарно и образуют периваскулярные сплетения, от которых отходят волокна, проникающие в стенку и распределяющиеся в адвентиции (t?nica ext?rna) и между последней и t?nica m?dia. Волокна иннервируют мышечные образования стенки, имея различную форму окончаний. В настоящее время доказано наличие рецепторов во всех кровеносных и лимфатических сосудах.
Первый нейрон афферентного пути сосудистой системы лежит в спинномозговых узлах или узлах вегетативных нервов (nn. spl?nchnici, n. v?gus); далее он идет в составе кондуктора интероцептивного анализатора (см. «Интероцептивный анализатор»). Сосудодвигательныи центр лежит в продолговатом мозге. К регуляции кровообращения имеют отношение gl?bus p?llidus, таламус, а также серый бугор. Высшие центры кровообращения, как и всех вегетативных функций, заложены в коре моторной зоны головного мозга (лобная доля), а также впереди и сзади нее. Корковый конец анализатора сосудистых функций располагается, по-видимому, во всех отделах коры. Нисходящие связи головного мозга со стволовыми и спинальными центрами осуществляются, по-видимому, пирамидными и экстрапирамидными трактами.
Замыкание рефлекторной дуги может происходить на всех уровнях центральной нервной системы, а также в узлах вегетативных сплетений (собственная вегетативная рефлекторная дуга).
Эфферентный путь вызывает вазомоторный эффект — расширение или сужение сосудов. Сосудосуживающие волокна проходят в составе симпатических нервов, сосудорасширяющие волокна идут в составе всех парасимпатических нервов краниального отдела вегетативной нервной системы (III. VII, IX, X), в составе передних корешков спинномозговых нервов (признается не всеми) и парасимпатических нервов сакрального отдела (nn. spl?nchnici pelv?ni).
ЕДИНСТВО ВЕГЕТАТИВНОЙ И АНИМАЛЬНОЙ ЧАСТЕЙ НЕРВНОЙ СИСТЕМЫ
Необходимо помнить, что вегетативная нервная система есть часть единой нервной системы. Поэтому в целом организме постоянно наблюдается сочетанная деятельность вегетативной и анимальной частей нервной системы с вовлечением центров, находящихся на различных уровнях нервной системы.
Рассмотрим такую сочетанную деятельность на примере регуляции акта мочеиспускания.
В едином акте мочеиспускания участвуют непроизвольные мышцы (m. detr?sor ves?cae и m. sphincter vesicae), иннервируемые вегетативными нервами, и произвольная (m. sph?ncter ur?thrae), иннервируемая анимальными. При этом вытеснитель мочи сокращается, а оба сфинктера расслабляются, хотя каждый из них иннервируется из разных частей нервной системы: сфинктер пузыря — из вегетативной, а сфинктер мочеиспускательного канала — из анимальной. Это происходит благодаря наличию общего центра координации в головном мозге, поскольку вегетативная и анимальная части составляют единую нервную систему.
В интеграции анимальной и вегетативной нервных систем большую роль играет лимбическая система.
Зоны Захарьина-Геда. В настоящее время имеются сведения об афферентной иннервации внутренностей анимальной нервной системой, что отмечалось выше. Возможно, этим объясняется давно известный симптом отраженных болей, наблюдаемый в клинике. Заболевания некоторых внутренних органов постоянно сопровождаются отраженными болями в определенных местах кожи. Таковы, например, боли в левой лопатке и левой руке при стенокардии, боли между лопатками при язве желудка, боли в правой подвздошной ямке при аппендиците и др. Эти боли локализуются в определенных кожных сегментах, соответствующих тем сегментам спинного мозга, куда поступают афферентные (чувствительные) волокна из пораженного внутреннего органа. Такие кожные сегменты, или зоны, называются зонами Захарьина — Геда по имени, описавших их авторов. Приводим схему этих зон (рис. 347).
Рис. 347.
Знание зон Захарьина — Геда помогает судить по болям в наружных покровах тела о состоянии органов внутри его полостей. Возможно, что зоны Захарьина-Геда обусловливают воздействие на внутренние органы применяемых в китайской медицине прижиганий или вкалываний игл (акупунктура) в определенные кожные точки.
Сегментарная иннервация органов. Внутриорганные нервы паренхиматозных органов распределяются, как и сосуды, соответственно сегментам органов. Так, в легких различают 10 нервных сегментов. В печени число их весьма варьирует и может достигать восьми. В почке отмечается 5 сегментов иннервации.






