Компоненты болевого синдрома
Уже давно отзвучали споры между представителями корковой и подкорковой теории боли. Огромный экспериментальный и теоретический материал, накопленный в лабораториях и клиниках, показал, что каждая теория содержит и правильные, и неправильные положения. Конечное болевое ощущение формируется как в коре, так и в подкорковых образованиях головного мозга. Но различные мозговые структуры вносят свой вклад, включая свои «тоны и обертоны» в ту сумму раздражений, ощущений, эмоций, физиологических, физико-химических и патологических процессов, из которых складывается комплексное чувство боли.
Исследователи шли разными путями к решению вопроса о формировании болевого ощущения в различных участках мозга. Одни раздражали электричеством отдельные ядра, ансамбли нервных клеток и изолированные нейроны; другие действовали на них химическими веществами; третьи разрушали нервные структуры; четвертые предпочитали электроэнцефалографические записи; пятые, наконец, вводили в организм определенные фармакологические препараты и на фоне их действия изучали болевое поведение. Проводились бесчисленные эксперименты на животных — крысах, кроликах, собаках, обезьянах, ставились исследования на добровольцах, изучались болевые синдромы в клиниках. Шагом вперед явилось применение метода изучения болевых реакций, разработанного на кафедре психофизиологии Парижского университета.
Наибольший интерес представляют опыты, в которых сочетались физиологические, электроэнцефалографические, биохимические, неврологические и гистологические исследования.
Читателю уже известно, что и в эксперименте, и в клинике широко применяется метод отведения электрических потенциалов от различных отделов головного мозга. При помощи специальной аппаратуры удается зарегистрировать на бумаге или фотопленке электрические токи, возникающие в мозгу. Запись этих токов — электроэнцефалограмма — представляет сложную кривую, отражающую электрическую активность огромного числа нервных клеток. На ней можно выделить волны различной величины, частоты, формы. Чаще всего это так называемый альфа-ритм (8—13 колебаний в 1 сек.), который можно обнаружить в коре и подкорковых образованиях головного мозга. При различных воздействиях на организм альфа-ритм ослабевает, и величина ритмичных колебаний уменьшается. Это явление называется реакцией активации пробуждения, десинхронизации. Как правило, оно наблюдается также и при болевых раздражениях и наиболее выражено (по нисходящему ряду) в теменных, затылочных, височных и лобных долях мозга. Однако реакция пробуждения не специфична для боли. И нельзя ее считать объективным проявлением чувства боли. Аналогичная картина возникает при зрительных, слуховых, температурных и даже тактильных раздражениях.
Развитие техники и электроники значительно расширило возможности электроэнцефалографического исследования деятельности головного и спинного мозга. Этому в немалой степени способствовало внедрение в практику физиологического и даже клинического эксперимента метода вживления электродов в различные отделы центральной нервной системы. Почти в любой физиологической лаборатории удается без особого труда записать электрическую активность зрительных бугров, подбугорья, ретикулярной формации, гиппокампа, миндалевидного ядра и т.д.
При незначительной боли, вызванной слабым током, когда животное настораживается, иногда делает прыжок и очень редко убегает, никаких особых изменений в электроэнцефалограмме отметить не удается. Но сильная боль, сопровождающаяся прыжком, бегством, криком и попыткой укусить или сорвать электроды, вызывает отчетливое изменение электрической активности в разных отделах мозга, особенно отчетливо выраженную в гиппокампе. В нем при боли регистрируются токи высокой амплитуды, ритмом 6—7 в 1 сек. Изменения электрической активности гиппокампа длятся обычно 8—14 сек. и постепенно затухают. Одновременно в коре головного мозга и ретикулярной формации развивается типичная реакция пробуждения. Однако ни в том, ни в другом отделе мозга не удается обнаружить какие-либо специфические для боли сдвиги.
Применение различных болеутоляющих веществ (морфина, петидина, декстроморамида, производных салициловой кислоты) в сочетании с болевым раздражением во многом изменяет течение физиологических реакций и характер электроэнцефалограммы. Под влиянием морфина исчезают два компонента болевой реакции — крик и попытка укусить электроды . При этом в гиппокампе отсутствуют характерные изменения электрической активности.
У крыс с разрушенной ретикулярной формацией исчезают другие компоненты болевого поведения — настораживание, прыжок, бегство. Разрушение миндалевидного ядра напоминает действие морфина. Разрушение передней части зрительных бугров и коры лобных долей мозга снимает конечную поведенческую реакцию животного — попытку укусить или сорвать электроды.
На Симпозиуме по проблеме боли в Париже весной 1967 г. была предпринята попытка подытожить наши представления о формировании болевого ощущения в головном мозгу. Было заслушано и обсуждено много докладов, высказано немало интересных мыслей, сделано не меньшее число различных предположений. Можно считать установленным, что ретикулярная формация регулирует и контролирует более примитивные, вызванные болевым раздражением реакции настораживания, прыжка и бегства. Возбуждение ретикулярной активирующей системы вовлекает в процесс другие граничащие с нею отделы мозга. Зрительные бугры, в частности их медиальные (внутренние) ядра, являются центром более сложных эмоциональных, аффективных реакций. Переднее ядро зрительных бугров передает информацию, поступающую с периферии в кору головного мозга, где происходит осознание боли и вступают в действие сложные механизмы приспособительных функций. В коре происходит отбор и оценка поступающих импульсов, здесь формируется целенаправленное болевое поведение, цель которого перестроить функции организма в условиях неотвратимого болевого ощущения.
Разрушение некоторых отделов зрительных бугров резко усиливает реакцию на боль. Даже слабое болевое раздражение сопровождается криком и стремлением укусить или сорвать электроды. В некоторых случаях крыса приходит при этом в состояние настоящего бешенства. Это подтверждает уже неоднократно высказанное предположение, что зрительные бугры располагают тормозными системами, смягчающими болевое ощущение. Разрушение этих систем резко усиливает эмоциональные проявления боли.
В конце 70-х годов группа исследователей, среди которых следует назвать уже известного нам американского физиолога Лима и французских ученых Альб-Фессар, Кротамера и Делакура, широко разрабатывает вопрос о мозговых системах, смягчающих (демпфирующих) болевые ощущения. >>>На Парижском симпозиуме по боли о них говорил в своем докладе французский невропатолог Гарсен, а на симпозиуме по боли, который проходил в рамках XXIV Международного физиологического конгресса в Вашингтоне, выступили сами авторы этих интересных исследований.
В эволюционном развитии мозга от низших форм живых существ к высшим возникла необходимость как-то заглушать или ослаблять сигналы о сверхсильных импульсах, поступающих в центральную нервную систему. В некоторых случаях необходимо было затормозить поток сигналов, в некоторых — изменить их характер (модальность). Это привело к возникновению в центральной нервной системе нисходящей (афферентной — от центра к периферии) импульсации, оказывающей свое влияние на глубоколежащие мозговые образования или на периферические нервные пути. В своих статьях и докладах Альб-Фессар рисует сложные пути нисходящих или, как она их называет, тормозных болевых систем. Они состоят из определенных участков коры головного мозга (чувствительных, двигательных, теменных, отчасти лобных) и некоторых подкорковых образований, в первую очередь так называемого хвостатого ядра. Раздражение определенных участков хвостатого ядра электрическим током подавляет неприятный, тягостный характер импульсов, поступающих в мозг. Американские хирурги Эрвин, Браун и Марк использовали это наблюдение в клинической практике. Им удалось облегчить тяжелые боли, раздражая хвостатое ядро через вживленные электроды. В последние годы (1970—1972) составлена карта распределения в мозгу точек, раздражение которых вызывает полнейшее обезболивание. Некоторые исследователи утверждают, что, стимулируя эти точки электрическим током, они могли на животных производить сложнейшие операции без всякого наркоза.
Существование смягчающих боль систем в головном мозгу во многих отношениях соответствует современным представлениям о формировании болевого ощущения. Если вспомнить теорию Гэда о протопатической и эпикритической чувствительности, о способности коры головного мозга переводить болевые ощущения в «подболевые», то и представление о нисходящих «противоболевых» системах как бы воплотится в плоть и кровь. Мельзак утверждает, что существует специальный контролирующий механизм в ретикулярной формации мозгового ствола, оказывающий постоянное тормозящее влияние на всю систему синаптических передач — с импульсов от периферии до спинного и головного мозга. Альб-Фессар, а за ней и Гарсен высказывают мысль, что теоретически можно снимать боль, раздражая нисходящие системы мозга, но рассматривают эту возможность как «несбыточную мечту». Не будем такими пессимистами! Развитие современной нейрохирургии и хирургии боли не оставляет сомнении, что подобные операции найдут, если уже не нашли, свое место в клинической практике.
Кора лобных долей мозга принимает участие в организации поведения, наиболее выгодного для организма в условиях болевого раздражения. Именно в коре лобных долей происходит превращение боли в страдание. Как здесь не вспомнить парадоксальные результаты лоботомии , когда больные продолжали испытывать боль, но не страдали от нее. Однако в коре происходит не только формирование чувства боли и выработка наиболее выгодных при данной ситуации поведенческих реакций. В ней имеются также механизмы, облегчающие и подавляющие боль. Здесь расположены центры эпикритической чувствительности, отсюда осуществляется регулирующее влияние на ретикулярную формацию.
Важнейшую роль в формировании болевого ощущения играют процессы возбуждения и торможения, протекающие в различных структурах головного мозга. Особенное значение имеют они, как показали классические исследования школы И. П. Павлова, для деятельности коры головного мозга.
Сочетание и взаимодействие возбудительного и тормозного процесса позволяют животному ориентироваться в сложных положениях и разбираться в потоке поступающих из внешнего мира разнообразных раздражений.
В борьбе организма с болевым ощущением торможение играет чрезвычайно важную, если не решающую роль. «Клетки больших полушарий,— говорит И. П. Павлов note 2 ,— в высшей степени чувствительны к малейшим колебаниям внешней среды и должны быть тщательно оберегаемы от перенапряжения, чтобы не дойти до органического разрушения. Таким ограничительным средством для клеток больших полушарий и является торможение».
Торможение дает клеткам мозга необходимый им отдых, способствует восстановлению их функций. Если раздражитель очень силен и превышает предел выносливости нервных клеток, он может привести последние к истощению и даже гибели. Эта угроза предотвращается своевременным развитием торможения, которое как бы ограждает нервные клетки от чересчур сильных воздействий, падающих на них из внешней или внутренней среды.
Несомненный интерес представляет в этом отношении так называемое запредельное, охранительное торможение, которое развивается при действии на организм чрезмерных раздражителей, даже условных.
Такие воздействия, как болезнь, перенапряжение, физическая боль, угрозы, психические потрясения и т.д., могут вызвать запредельное торможение (в особенности, если они действуют продолжительное время).
При запредельном торможении нарушается правило зависимости эффекта от величины и интенсивности раздражителя и сильные раздражители начинают действовать слабев умеренных. Это объясняется тем, что нервные клетки защищаются от истощения и разрушения при помощи широко распространяющегося процесса торможения.
Повседневная жизнь дает немало примеров стойкого запредельного торможения. Все мы из личного опыта знаем, какую острую, нестерпимую боль вызывают сравнительно небольшие нарушения целости тканей и как спокойно переносятся подчас чрезмерно сильные болевые раздражения. Торможение играет в этих случаях не только роль защитного фактора организма, но и своеобразного терапевтического средства.
Таковы отдельные этапы формирования единого, комплексного болевого ощущения, которое заставляет нас страдать, требовать помощи, стонать или кричать, иногда плакать. С большей или меньшей вероятностью мы подошли к пониманию его механизмов. Но анализ не будет полным, если мы пройдем мимо важнейшего — вегетативного отдела нервной системы, который не только выполняет приказы, поступающие из ее высших отделов, но и сам в значительной мере определяет состояние механизмов, регулирующих всю жизнедеятельность организма.
В специальной литературе это носит название «регуляция регуляторов». Вегетативная нервная система с ее центральными и периферическими образованиями, гуморальными и гормональными составными частями принимает самое энергичное участие во всех болевых процессах, организуя не только цепь болевых реакций, но и определяя характер самого болевого ощущения.
В мозгу, как мы уже знаем, существуют сложные вегетативные системы, которые приходят в деятельное состояние под влиянием медиаторов — ацетилхолина, адреналина или серотонина. Они и получили название холинергических, адренергических и серотонинергических структур. От их состояния в немалой степени зависит формирование болевого ощущения.
Введение в организм эзерина — вещества, подавляющего фермент холинэстеразу и тем самым усиливающего действие ацетилхолина,— изменяет в опытах на крысах соотношение компонентов болевого синдрома. Реакция бегства у крысы становится слабее, реакция крика и кусания электродов усиливается. Болевое возбуждение обостряется и удлиняется. Противоположное действие отмечается при введении холинолитического препарата — атропина. Стремление к бегству усилено, крик и кусание электродов ослаблены.
Однако немецкий ученый Герц, используя другие холинергические препараты (ареколин, треморин), пришел к выводу, что они ослабляют болевое ощущение и одновременно подавляют условно-рефлекторную деятельность крыс, т.е. действуют через высшие отделы головного мозга.
Шарпантье считает, что холинергические вещества осуществляют свое действие, возбуждая гиппокамп, а адренергические — ретикулярную формацию.
Советский фармаколог П. П. Денисенко также установил, что холин– и адренергические элементы лимбической системы, ретикулярной формации среднего мозга и подбугорья играют важную роль в формировании болевых реакций, причем, по его данным, ведущее значение имеют М –холинергические структуры. Ему удавалось во много раз усилить обезболивающее действие морфина, вводя его одновременно с препаратами, снижающими активность центральных М –холинергических образований. Однако следует признать, что вопрос о взаимоотношении холин– и адренергических систем в мозгу при формировании болевого ощущения далеко еще не ясен. Возможно, что у разных видов животных, а тем более у человека, роль их различна.
Еще менее ясен вопрос об участии серотонина в возникновении болевого ощущения. При полном освобождении головного мозга от серотонина или, напротив, при значительном увеличении его содержания в ткани мозга (то и другое легко осуществить с помощью определенных фармакологических препаратов) порог болевого ощущения резко изменяется, но подчас в одну и ту же сторону. По всей вероятности, серотонин контролирует в первую очередь эмоциональные реакции, обусловленные болью, и в гораздо меньшей степени затрагивает поведение животного, например реакцию кусания электродов .
Таким образом, можно предположить, что в центральной нервной системе существует два этажа (или, как говорят физиологи, два уровня), объединяющие болевые раздражения и превращающие их в чувство боли. На первом формируется состояние общей настороженности животного. Это происходит в ретикулярной формации и осуществляется ее адренергическими элементами. Здесь начало простых неспецифических ответов на боль, выражающихся в прыжке и бегстве. Возбуждение ретикулярной формации активирует кору головного мозга и вызывает в ней феномен пробуждения, т.е. подавление альфа-ритма.
На втором этапе, в лимбической системе (гиппокампе, миндалевидном ядре) и зрительных буграх развиваются специфические болевые реакции. Это ведет к возникновению эмоциональных, аффективных состояний. С этим уровнем у крысы связаны реакции крика и кусания электродов. Здесь действует холинергическая система. Центром ее является гиппокамп. В нее же входят некоторые участки зрительных бугров, но уже не возбуждающие, а тормозящие болевое ощущение.
Оба болевых уровня находятся под неослабным контролем коры головного мозга. Искусственное разрушение коры полностью снижает осмысленную реакцию, наблюдающуюся при болевом раздражении,— попытку укусить электроды, но не подавляет крика. Крик — прерогатива подкорки!
У человека кора превращает чувство боли в страдание, т.е. облекает болевой рефлекс в психическую оболочку. Переход боли в страдание необязателен, но, если он наступил, вся регуляция функций может оказаться под угрозой. Информация, поступающая в высшие отделы мозга, становится неточной в одних случаях, избыточной в других. Хаотическое пароксизмальное вмешательство высших нервных центров в физиологические процессы приводит к дезорганизации гомеостатических (приспособительных) механизмов, к возникновению невротических состояний. Цепь болевых реакций превращается в боль-болезнь.
Еще несколько лет тому назад на этом можно было бы поставить точку. Но за последние годы широкое звучание получила теория канадских ученых Мельзака и Уолла, известная в нашей стране под названием «теории входных ворот», или, точнее, «теории контроля афферентного входа». Мимо нее в настоящее время не проходит ни один исследователь, занимающийся проблемой боли. И в докладах, и в дискуссиях на симпозиумах 1973 г. в Ленинграде и Вашингтоне — многие неясные и нерешенные проблемы физиологии и, особенно, фармакологии боли рассматривались с точки зрения Мельзака.
По-видимому, теории интенсивности и специфичности не могут полностью расшифровать противоречивые аспекты болевого ощущения. Многие физиологи и фармакологи считают, что множество фактов, накопленных исследователями в эксперименте и клинике, не позволяют считать, что боль является специфической модальностью со своим исключительно обособленным рецепторным аппаратом, специальными путями проведения и центральными механизмами. В то же время и сторонники теории специфичности приводят множество неоспоримых фактов, подтверждающих существование анатомо-физиологической системы боли. Возможно, что правы и те и другие. Попыткой найти какую-то среднюю линию и является теория «входных ворот».
Мельзак и его сотрудники утверждают, что формирование болевого ощущения начинается в задних рогах спинного мозга. Здесь находятся нервные клетки, модулирующие импульсы, поступающие с периферии. Решающую роль в превращении или непревращении поступившего сигнала в болевой играет желатинозная субстанция . Она как бы «настраивает» клетки задних рогов, задает тон, включает определенные тоны и обертоны. Импульсы от рецепторов приходят в спинной мозг по толстым (А –бета) и тонким (А –дельта и С) волокнам, но проникают они в задние рога через «ворота» желатинозной субстанции. Одновременно нервные клетки задних рогов получают из вышележащих нервных центров нисходящие импульсы, аналогичные описанным выше «противоболевым», тормозным сигналам. На рис. 22 представлена схема Мельзака и Уолла, облегчающая понимание сложных процессов, разыгрывающихся у «ворот» центральной нервной системы.
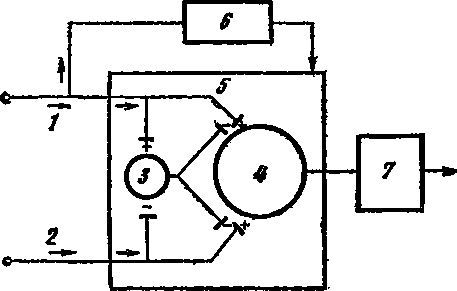
Рис. 22. Схематическая диаграмма теории «входных ворот» (по Мельзаку и Уоллу, 1965)
1 — толстое миелиновое волокно; 2 — тонкое безмиелиновое волокно; 3 — желатинозная субстанция; 4 — клетки центральной нервной системы, получающие первые афферентные сигналы; 5 — контроль, осуществляемый «входными воротами»; 6 — контроль, осуществляемый высшими отделами центральной нервной системы; 7 — система действия. Возбуждение +, торможение —. Торможение, которое оказывает желатинозная субстанция на клетки центральной нервной системы, усиливается импульсацией с толстых нервных волокон и ослабевает при импульсации с тонких нервных волокон. Центральные образования, контролирующие «входные ворота», возбуждаются под влиянием сигналов, поступающих по толстым нервным волокнам. Клетки центральной нервной системы стимулируют систему действия
Итак, клетки спинного мозга — так называемые передаточные клетки (ПК) — получают импульсы как по толстым, так и тонким нервным волокнам (1, 2). Импульсы модулируются желатинозной субстанцией (ЖС). Под ее влиянием они могут усиливаться (+) и ослабевать (—). Сигналы с толстых волокон подавляют передачу возбуждения (закрывают ворота), с тонких — активируют ее (открывают ворота). Импульс, поступивший по толстым волокнам («быстрая боль»), мгновенно оценивается высшими уровнями нервной системы. Эту оценку приводит специальная контролирующая система головного мозга. В тот момент, когда возбуждение в передаточных клетках спинного мозга достигло какого-то критического уровня, «входные ворота» открываются и происходит формирование болевого ощущения со всеми его болевыми и противоболевыми компонентами. При этом желатинозная субстанция действует в двух направлениях. Она способна блокировать импульсы, поступающие с периферии, и может способствовать образованию медиаторов в нервных клетках, тем самым усиливая или притормаживая проведение возбуждения в центральной нервной системе.
В дальнейшем Мельзак предпринял попытку расчленить единое болевое ощущение на три основных слагаемых. Физиологические особенности боли как одного из чувств (шестое чувство) регулируются спинно-бугровой системой, передающей импульсы с периферии в мозг. Мощная мотивационно-эмоциональная сфера боли связана с состоянием лимбико-ретикулярных структур мозга, которые побуждают живой организм совершать те или иные действия. И, наконец, познание, оценка боли и сопоставление ее с накопленным опытом совершается высшими отделами головного мозга, в первую очередь корой больших полушарий.
Таким образом, сигналы, поступающие от рецепторов, иногда достаточно сильные, чтобы вызвать боль, демпфируются и не доходят до сознания. В этом «повинна» желатинозная субстанция — либо вся, либо отдельные ее слои (пластины). Она придает им определенную модальность, которая может быть воспринята корой мозга и как ощущение прикосновения, и как чувство боли. В свою очередь сама центральная нервная система может затормозить превращение импульса в болевой, вводя в действие нисходящие противоболевые механизмы. Все это приводит к задержке иногда сильного, иногда даже сверхсильного импульса у «входных ворот» и к отсутствию болевого ощущения.
Теория Мельзака позволяет весьма убедительно объяснить действие некоторых противоболевых препаратов на организм. Быть может, поэтому она получила столь широкое распространение среди фармакологов, занимающихся проблемой обезболивания.
Теперь, как в любом математическом расчете, можно подвести некоторые итоги и подсчитать суммы прибылей и убытков. Формирование болевого ощущения — сложный, многозвеньевой процесс, охватывающий ряд анатомических и физиологических систем. Чувство боли возникает в результате системной деятельности нервной системы, т. е. вовлечения в действие самых различных ее формаций, ансамблей рецепторов, нейронов, секреторных клеток, периферических и центральных проводящих путей, различных как по строению, так и особенностями регулирования и химической передачи возбуждения. Функциональная система, реализующая болевое ощущение, включает и «болевые», и «противоболевые» механизмы, и смягчающие, и обостряющие боль настроечные компоненты, и пусковые, и выключающие элементы, Как и всякая функциональная система, болевая система «является конкретным физиологическим аппаратом, благодаря которому осуществляются саморегуляция и гомеостаз» note 3 .
Все эти факты и обобщения необходимо учитывать в борьбе с болью. Разумеется, не каждый лабораторный или клинический эксперимент может быть использован в медицинской практике. Но некоторые выводы из теоретических положений должны быть учтены и теоретиками, и врачами.