Оплодотворение
Оплодотворением называется процесс слияния зрелых мужской (сперматозоид) и женской (яйцеклетка) половых клеток, в результате чего возникает зигота, несущая генетическую информацию как отца, так и матери.
Процесс созревания мужских и женских половых клеток очень сложен. Сперматогенез совершается в извитых семенных канальцах мужских гонад. Он завершается в период половой зрелости образованием зрелых сперматозоидов, обладающих способностью к оплодотворению. Полному созреванию предшествует процесс редукционного деления, в результате которого в ядре сперматозоида содержится гаплоидный набор хромосом.
Сперматозоиды бывают двух видов: носители половых Хи Y-хромосом. При слиянии с яйцеклеткой сперматозоида, являющегося носителем половой Х-хромосомы, из образующейся зиготы развивается эмбрион женского пола, при слиянии сперматозоида, имеющего половую Y-хромосому, возникает зародыш мужского пола (яйцеклетка всегда является носительницей половой Х-хромосомы).
Зрелый сперматозоид имеет длину до 50-60 мкм и состоит из головки, шейки и хвостовой части. Головка сперматозоида, имеющая овальную форму, содержит ядро, окруженное тонким слоем протоплазмы. Шейка имеет протоплазму, содержащую видоизмененную центросому, которая способствует процессу дробления оплодотворенной яйцеклетки. Хвостик состоит из протоплазмы и выполняет двигательные функции, В результате колебательных движений хвостовой части сперматозоиды способны совершать самостоятельные движения в половом тракте женщины со скоростью 2-3 мм/мин. Способность к движениям сперматозоиды получают после их контакта с секретом семенных пузырьков и предстательной железы. Такая смесь носит название семенной жидкости, или спермы. При половом сношении во влагалище женщины изливается в среднем около 3-5 мл спермы, в которой содержится 300-500 млн сперматозоидов. Использование электронной микроскопии позволило установить весьма сложную структуру сперматозоида (рис. 3.1).
Семенная жидкость имеет сложный состав и содержит фруктозу, белковые вещества, протеазы, кислую фосфатазу, лимонную кислоту и биологически активные вещества – простагландины. Семенная жидкость обладает выраженной антигенной активностью, при этом антигены обнаружены как в самих сперматозоидах, так и в жидкой части спермы. Полагают, что число антигенных компонентов в семенной жидкости достигает 12. В процессе оплодотворения происходит нейтрализация антигенов сперматозоидов.
Развитие яйцектеток связано с ростом и развитием первичных фолликулов, находящихся в корковом слое яичников. Созревание яйцеклетки условно можно разделить на созревание ядра и созревание цитоплазмы. Под созреванием ядра понимают совокупность процессов, переводящих ядро из стадии динлотена I (или стадии герминативного пузырька) до метафазы второго мейотического деления. Созревание ядра не включает в себя завершение мейоза, так как только проникновение сперматозоида внутрь яйцеклетки вызывает завершение второго редукционного деления.
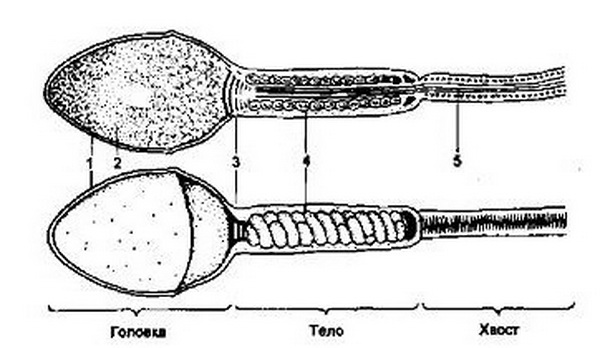
Рис .3.1. Строение сперматозоида человека (электронно-микроскопическая схема), 1 – акросома; 2 – ядро; 3 – шейка; 4 – митохондрии; 5 – осевые нити.
От периода наступления полового созревания до менопаузы у женщины в каждом менструальном цикле обычно созревает одна яйцеклетка. Овоцит первого порядка пре-вращается в овоцит второго порядка при отщеплении первого полярного тельца. В мо-мент овуляции овоцит второго порядка оказывается блокированным на стадии метафазы второго мейотического деления. Созревание яйцеклеток стало известно в деталях благодаря возможности культивирования яйцеклеток in vitro, что в настоящее время широко используют при экстракорпоральном оплодотворении.
Зрелая яйцеклетка состоит из ядра, цитоплазмы, окружена блестящей оболочкой и клетками лучистого венца, представляющего собой остатки гранулезных клеток фоллику-ла (рис. 3.2). Женская половая клетка, как и мужская, обладает антигенными свойствами. Особенно богата различными антигенами ее блестящая оболочка.
Прежде чем анализировать механизмы оплодотворения, т.е. процесс слияния яйцеклетки со сперматозоидом, следует осветить вопросы транспорта гамет. В первую очередь это касается продвижения сперматозоидов по каналам репродуктивной системы женщины. При нормальной эякуляции во влагалище попадает в среднем около 100 млн сперматозоидов, некоторые из которых имеют те или иные морфологические либо функциональные отклонения. Часть сперматозоидов, в том числе и неполноценных, остается во влагалище и подвергается фагоцитозу, Вместе со сперматозоидами во влагалище попадают и другие составные части спермы, при этом особая роль принадлежит проста гланд инам. Под их влиянием происходит активация сократительной активности матки и маточных труб, что очень важно для нормального транспорта гамет.

Рис. 3.2. Зрелая яйцеклетка, окру-женная клетками лучистого венца.
Из влагалища, имеющего у здоровой женщины кислую среду (кислая среда неблагоприятна для жизнедеятельности сперматозоидов), последние быстро поступают в цервикальную слизь, которая во время полового акта под влиянием сокращений мышц шейки матки выделяется из цервикального канала. Наличие слабощелочной реакции цервикальной слизи способствует повышению двигательной активности сперматозоидов.
Оптимальный состав цервикальной слизи формируется к моменту овуляции в основном под влиянием эстрогенных гормонов яичников. В этот период мицелии слизи располагаются в виде своеобразных цепочек, ориентированных по силовым линиям магнитного поля земли. Сперматозоиды продвигаются по мицелиям слизи по направлению к матке, но лишь при определенной архитектонике цепочек мицелл. Турбулентные движения сперматозоидов наиболее выражены в пристеночных областях шейки матки, при этом часть сперматозоидов на некоторое время может депонироваться в криптах шейки матки, создавая тем самым своеобразный резерв спермы, откуда в дальнейшем может происходить дополнительное их поступление вверх по каналам репродуктивной системы.
В верхних отделах полового тракта женщины начинается процесс, называемый капацитацией спермы, – приобретение ею благодаря сложным изменениям способности к оплодотворению. В результате капацитации сперматозоиды приобретают способность к акросомальной реакции. Помимо этого, капацитация выражается в изменениях движений хвостовых частей сперматозоидов (наличие сверхактивной подвижности). Тонкие механизмы капацитации до настоящего времени полностью не изучены. Время капацитации различно у разных сперматозоидов, что, по-видимому, является важной приспособительной реакцией для процесса оплодотворения. Капацитированные сперматозоиды очень активны, однако продолжительность их жизни меньше, чем некапацитированных. Капацитированные сперматозоиды обладают повышенной способностью пенетрировать ткани, что имеет решающее значение в процессе оплодотворения яйцеклетки.
Транспорт сперматозоидов в матку, а затем и в маточные трубы в основном обеспечивается сокращениями гладкой мускулатуры этих органов. Полагают также, что трубно-маточные сфинктеры являются своеобразными дозаторами поступления сперматозоидов из полости матки в просветы маточных труб.
Наряду с сократительной способностью маточных труб, которые находятся под сложным гормональным воздействием (эстрогены, андрогены, окситоцин), а также под влиянием простагландинов, большое значение в продвижении сперматозоидов, помимо их собственной высокой кинетической активности, принадлежит таким факторам, как движения микроворсин реснитчатого эпителия эндоцервикса и ток жидкости в просвете маточной трубы. Таким образом, перемещение сперматозоидов по каналам репродук-тивной системы женщины представляет собой чрезвычайно сложный многокомпонентный процесс.
Результаты современных научных исследований свидетельствуют о наличии двухфазности процесса транспорта сперматозоидов по маточным трубам. В первую (короткую) фазу сперматозоиды начинают быстро поступать ампулу трубы. Эта фаза длится всего несколько минут и регулируется сократительной активностью матки и маточных труб. Данная фаза сменяется более длительной второй фазой, в течение которой сперматозоиды со значительно меньшей скоростью транспортируются к месту оплодотворения.
В ампулярной части маточной трубы нормальное количество сперматозоидов в длительной фазе транспорта сохраняется на определенном уровне благодаря непрерывному возмещению уходящих в брюшную полость сперматозоидов за счет тех половых клеток, которые депонированы в нижних отделах полового тракта (крипты шейки матки и др.).
Большое значение в транспорте гамет и оплодотворении имеет так называемый захват яйцеклетки ампулярным отделом маточной трубы. Этот процесс заключается в том, что ампулярный отдел трубы своими фимбриями как бы накрывает яичник в том месте, где произошла овуляция; при этом значительно облегчается перемещение зрелой яйцеклетки в ампулу маточной трубы. Механизм этого феномена окончательно еще не исследован.
Существенным является вопрос о выживаемости сперматозоидов в половых путях женщины. Некоторые авторы полагают, что жизнеспособность спермы сохраняется в течение нескольких (до 5) дней. Однако следует учитывать, что сохранение подвижности сперматозоидов не обязательно свидетельствует об их оплодотворяющей способности.
А в наиболее благоприятных условиях, когда мужские половые клетки находятся в цервикальной слизи на фоне высокого содержания эстрогенов в организме, оплодотворяющая способность сперматозоидов держится до 2 сут после эякуляции во влагалище. В связи с этим принято считать, что для достижения беременности оптимальная частота половых сношений в период до и после овуляции должна быть каждые 2 дня. При более частых половых сношениях фертильность спермы снижается.
Перед оплодотворением зрелая яйцеклетка со всех сторон окружена лучистым венцом (corona radiata). Отчетливо заметна блестящая оболочка (zona pellucida). Уже через несколько минут после разрыва фолликула (овуляции) яйцеклетка попадает в полость маточной трубы. Этому процессу способствует ряд факторов: "захват" ее фимбриями маточной трубы со стороны яичника, в котором произошла овуляция, направление тока фолликулярной жидкости при разрыве фолликула и др. Этим факторам принадлежит очень важная роль в первоначальном транспорте яйцеклетки, которая лишена самостоятельной подвижности. Способность яйцеклетки к оплодотворению в среднем составляет 24 ч. Современные методы диагностики (ультразвуковое исследование, лапароскопия) позволяют не только наблюдать за процессом овуляции, но и фиксировать этот процесс на фотопленку.
Яйцеклетка, попавшая в ампулярный отдел маточной трубы, быстро окружается большим количеством сперматозоидов, являющихся носителями как Х-, так и Y-половых хромосом. Под микроскопом сперматозоиды с Х-хромосомой имеют несколько более крупные размеры, чем несущие Y-хромосому. Сперматозоиды, окружающие яйцеклетку, начинают пенегрировать в клетки лучистого венца. Процесс пенетрации обусловлен наличием ряда ферментов, которые содержатся как в головке сперматозоида, так и в трубной жидкости (рис. 3.3).
Сразу же после слияния мембран половых клеток происходит кортикальная реакция яйцеклетки, являющаяся составной частью обеспечения блока полиспермии. После этого хромосомы зиготы вступают в первое митотическое деление, которое наступает через 24 ч после начала оплодотворения. Ядро оплодотворенной яйцеклетки (зиготы) содержит диплоидный набор хромосом (46). Таким образом, новый организм является носителем генетической информации обоих родителей.
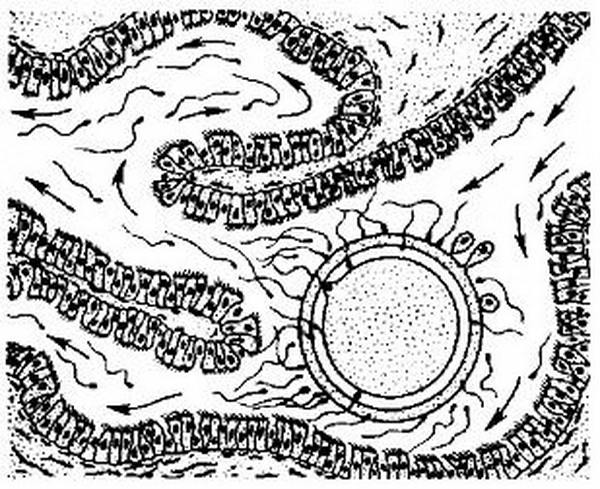
Рис. 3.3. Пенетрация сперматозоидов в яйцеклетку в ампулярном отделе маточной трубы (схема).
После оплодотворения (через 24 ч) начинается дробление оплодотворенной яйцеклетки. Первоначально дробление имеет синхронный характер. Через 12 ч от начала возникновения 2 бластомеров возникают 4 бластомера и т.д. К 96 ч от момента слияния ядра сперматозоида с ядром яйцеклетки зародыш состоит из 16-32 бластомеров (стадия морулы). На этой стадии оплодотворенное яйцо (зигота) попадает в матку.
Поскольку дробящаяся яйцеклетка не обладает самостоятельной подвижностью, ее транспорт определяется взаимодействием сократительной активности маточной трубы (основной фактор), движениями цилиарного эпителия эндосальпинкса и капиллярным током жидкости в направлении от ампулярного конца маточной трубы к матке.
Транспорт яйцеклетки по маточной трубе находится под воздействием гормонов. Как известно, после овуляции на месте лопнувшего фолликула образуется новая эндокринная железа – желтое тело. Оно выделяет как прогестерон, так и эстрогенные гормоны. Именно этим двум половым гормонам принадлежит ведущая роль в обеспечении кинетики маточных труб. Под влиянием относительно низкого содержания прогестерона и более высокой концентрации в крови эстрогенов (что имеет место непосредственно после овуляции) повышается тонус ампулярно-перешеечного отдела трубы. В результате яйцеклетка задерживается в ампулярном отделе, где происходит ее оплодотворение и начинается процесс клеточного деления с образованием бластомеров. В дальнейшем происходит постепенное продвижение оплодотворенной яйцеклетки по перешейку маточной трубы к матке. Под влиянием нарастающих концентраций прогестерона желтого тела сократительная функция маточных труб приобретает перистальтический характер, при этом волны сокращений направлены в сторону матки. Происходит расслабление трубно-маточного соединения, и яйцеклетка из маточной трубы попадает в полость матки.
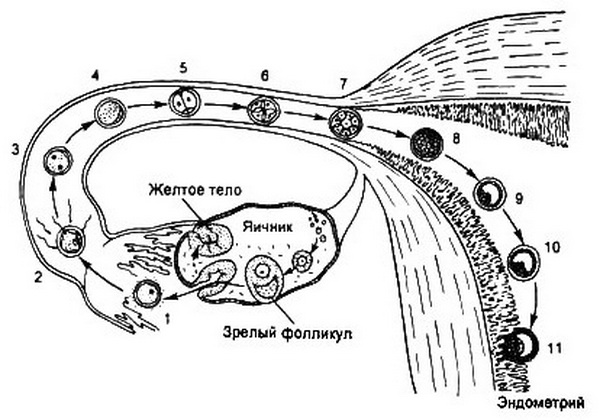
Рис. 3.4. Транспорт оплодотворенной яйцеклетки по маточной трубе вплоть до имплантации (схема).
1 – яйцеклетка в ампуле маточной трубы; 2 – оплодотворение; 3-7 – различные стадии образования бластомеров; 8 – морула; 9, 10 – бластоциста; И – импланта-ция.
Анализируя механизмы гормональной регуляции кинетики маточных труб, следует подчеркнуть, что только определенное соотношение между концентрациями в крови прогестерона и эстрогенов обеспечивает описанное выше четкое физиологическое взаимодействие различных отделов маточной трубы и своевременный транспорт плодного яйца в полость матки. Наряду с прогестероном и эстрогенами в осуществлении физиологической кинетики маточных труб в ранние сроки беременности известное значение отводится таким гормонам, как тестостерон и кортизол. Однако роль этих гормонов изучена в настоящее время еще недостаточно.






