Причины наступления родов
Причины наступления родовой деятельности до настоящего времени не установлены.
Теории причин развития родовой деятельности. Для объяснения причин наступления родовой деятельности было выдвинуто немало теорий, многие из которых представляют исторический интерес.
По мнению Гиппократа, роды наступают потому, что плод за счет голода, наступившего в конце беременности, сам выходит из полости матки, упираясь ножками в ее дно (головкой вперед). Согласно теории "инородного тела", роды наступают потому, что нарушается интимная связь между маткой и плодом за счет жирового перерождения тканей плаценты и эндометрия.
В последующем были выдвинуты различные теории наступления родов (механическая, иммунная, плацентарная, химическая, гормональная, эндокринная) .
Сторонники механической теории считали, что причинами возникновения родов являются возбуждение нервных рецепторов, расположенных в нижнем сегменте матки, в результате давления головкой плода.
С развитием учения об иммунитете связано появление иммунной теории, согласно которой в ответ на выделение синцитиотоксинов в
В настоящее время, помимо самопроизвольных родов, выделяют:
1. индуцированные роды – искусственное родовозбуждение по показаниям со стороны либо матери, либо плода;
2. программированные роды предусматривают процесс рождения ребенка в дневное, удобное для врача время. С этой целью в срок предполагаемых родов при полной зрелости плода проводится родовозбуждение.
В плаценте вырабатываются антитела – синцитиолизины. К концу беременности количество синцитиотоксинов возрастает настолько, что не происходит их нейтрализации. За счет этого матка становится легковозбудимой, в ней возникают импульсы, необходимые для сокращения. Однако поиски специфических синцитиотоксинов не увенчались успехом. Возможен другой механизм влияния иммунологических реакций на развитие родовой деятельности. По мнению некоторых ученых, рождение плода может уподобляться реакции отторжения трансплантата.
Согласно плацентарной теории, роды начинаются с возникновением различных изменений в плаценте, которые заключаются в перерождении ворсин и прекращении тормозящего влияния трофобласта на сократительную деятельность матки, а также с появлением в плаценте веществ, вызывающих схватки и начало родового акта. Эта теория не получила подтверждения.
Согласно химической теории, начало родового акта сопряжено с изменением состава неорганических веществ в матке и ионной среды в организме беременной, с накоплением угольной кислоты и других веществ, вызывающих сокращение мышц.
С развитием эндокринологии причины наступления родов стали объяснять изменением гормонального фона перед родами, способствующими повышению сократительной способности матки (эндокринная теория).
Представленный перечень теорий наступления родовой деятельности указывает на сложность изменений, происходящих у беременных перед родами и в родах.
Современные представления о причинах наступления родов. Роды протекают при наличии сформированной "родовой доминанты", представляющей собой единую динамическую систему, объединяющую как высшие центры регуляции (центральная и вегетативная нервная система, гормональная регуляция), так и исполнительные органы (матка и фетоплацентарный комплекс) (рис. 5.1).
Нормальный родовой акт определяется вовлечением в доминантный процесс коры большого мозга, в частности височных долей больших полушарий, а также значительным увеличением межполушарных связей, облегчающих координацию соматических функций.
Важная часть координационных центров родовой деятельности находится в подкорковых структурах мозга: в гипоталамусе – в миндалевидных ядрах лимбического комплекса, гипофизе.
Перед началом родов постепенно усиливаются тормозные процессы в коре большого мозга и повышается возбудимость подкорковых структур, регулирующих родовую деятельность. Одновременно увеличивается возбудимость периферических отделов нервной системы, в частности интерорецепторов, передающих возбуждение с половых органов. Афферентная импульсация от матки, которая формирует безусловные рефлексы, связанные с родовым актом, усиливается за счет зрелости фетоплацентарного комплекса. Перед родами она превышает порог чувствительности воспринимающих подкорковых структур, способствуя наступлению родов.
Выраженность рефлекторных реакций, лежащих в основе родов, зависит от тонуса различных отделов вегетативной нервной системы, иннервируюЩих матку. Все отделы матки (тело, нижний сегмент) имеют двойную вегетативную иннервацию. Адренергическая иннервация преобладает в продольно расположенных мышечных пучках в теле матки. Холинергическая иннервация наблюдается главным образом в циркулярно и спиралеобразно расположенных мышечных волокнах, находящихся преимущественно в нижнем сегменте матки. Там же расположены М-холинорецепторы. Адренорецепторы в матке представлены двумя типами: cqи сЛ-адренорецепторами. Они расположены на мембране гладкой мышечной клетки. арАдренорецепторы вызывают повышение возбудимости, тонуса и сократительной активности миометрия; В2-адренорецепторы оказывают противоположное действие на миометрии. Воздействие на В2-адренорецепторы вызывает снижение тонуса, возбудимости и сократительной активности матки. Перед родами увеличиваются количество и активность арадренорецепторов и Мхолинорецепторов.
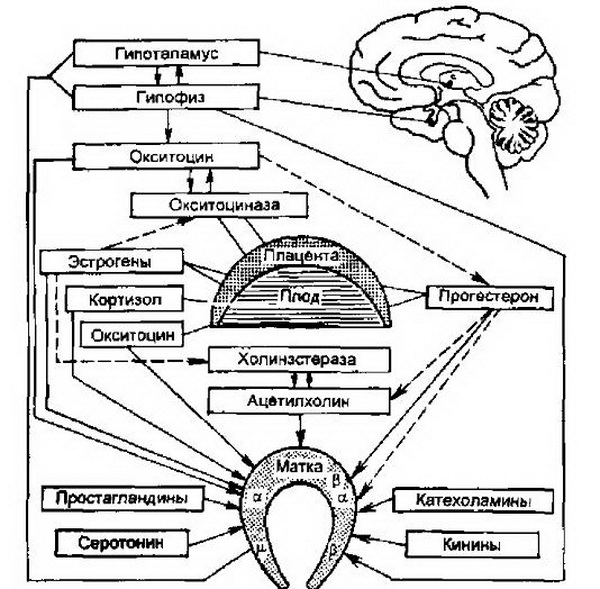
Рис. 5.1. Регуляция сократительной деятельности матки (схема). Сплошные стрелки – активация (стимуляция), пунктирные – угнетение (подавление): а – а-адренорецепторы; р – (3-адренорецепторы; р – М-холинорецепторы.
Вегетативная регуляция сокращений матки осуществляется через медиаторы, главными из которых являются ацетилхолин, адреналин и норадреналин.
Ацетилхолин оказывает стимулирующий эффект на мышцы матки.
Перед родами и в родах в плазме крови беременных наблюдается высокий уровень активной формы ацетилхолина при одновременно низкой активности ацетилхолинэстеразы.
Катехоламины (адреналин, норадреналин) опосредуют адренергические влияния на миометрий, взаимодействуя с аг и р2-адренорецепторами гладких мышечных клеток. Активирующий эффект катехоламинов в основном реализуется их воздействием на арадренорецепторы гладких мышечных клеток миометрия. Тормозное влияние катехоламинов на миометрий связано с взаимодействием их с рЛ-адренорецепторами гладких мышечных клеток.
Особенности нервной регуляции родовой деятельности сопряжены с изменением гормонального статуса перед родами. Только при наличии определенных гормональных соотношений возможны рефлекторная возбудимость матки и те нейрофизиологические изменения, при которых она способна регулярно сокращаться на протяжении родов. Из всего комплекса регулирующих компонентов, дублирующих друг друга в период подготовки к родам, особое значение имеет повышение синтеза эстрогенов (в основном активной их фракции – эстрадиола) на фоне снижения уровня основного гормона беременности – прогестерона, блокирующего сокращение мышц матки. Перед родами содержание прогестерона и его метаболитов в крови и моче снижается и соотношение в моче эстриол/прегнандиол составляет 1:1 (во время беременности 1:100).
Под действием эстрогенов происходят следующие изменения:
1. увеличивается кровоток в миометрий, интенсивность окислительновосстановительных процессов, синтез сократительных белков миометрия (актомиозин), энергетических соединений (АТФ, гликоген) и утеротонических простагландинов;
2. повышается проницаемость клеточных мембран для ионов (К , Са , Na ), приводящих к снижению мембранного потенциала покоя, увеличению чувствительности клеток миометрия к раздражению;
3. депонируется кальций в саркоплазматической сети;
4. повышается активность фосфолипаз и скорость "арахидонового каскада" с образованием простагландинов.
Подобные изменения способствуют интенсификации сократительной способности матки, ускорению "созревания" ее шейки.
В развитии родовой деятельности, кроме эстрогенов, ключевую роль играют простагландины , которые, по современным представлениям, являются основными стимуляторами начаЛ родовой деятельности. Местом синтеза простагландинов в беременной матке являются плодные (амнион и хорион) и децидуальная оболочки. При этом в амнионе и хорионе образуется простагландин Е (ПГЕ), а в децидуальной оболочке и миометрий синтезируется как ПГЕ, так и PGF2a (материнские). Выработка простагландинов в конце беременности обусловлена процессами старения и дегенерации структурных элементов плаценты, децидуальной оболочки, амниона, поскольку эти процессы сопряжены с активацией фосфолипаз и образованием арахидоновой кислоты, а в последующем – простагландинов. Простагландины стимулируют следующие процессы:
1. образование на мембране а-адренорецепторов и рецепторов к другим утеротоническим соединениям (ацетилхолин, окситоцин, серотонин);
2. обеспечение автоматического сокращения матки (схватки);
3. угнетение продукции окситоциназы.
Наряду с повышением синтеза эстрогенов и простагландинов для начала родов имеет важное значение повышение активности других нейрогуморальных медиаторов и гормонов (окситоцин, серотонин, кинин, гистамин).
Окситоцин является важным регулятором сократительной деятельности матки. Многими авторами он рассматривается как пусковой фактор в развитии родовой деятельности. Окситоцин – это синергист ацетилхолина и простагландинов. Его концентрация в плазме крови возрастает по мере увеличения срока беременности. Действие окситоцина на сократительную деятельность матки зависит от гормонального фона, прежде всего от оптимального уровня эстрогенов, а также функционального состояния матки, ее готовности к спонтанной активности.
Действие окситоцина связано со следующими процессами:
1. усиление мембранного потенциала и повышение возбудимости мышечной клетки;
2. увеличение скорости связывания ацетилхолина рецепторами миометрия и освобождение его из связанного состояния;
3. возбуждение а-адренорецепторов;
4. угнетение активности холинэстеразы.
В результате действия окситоцина повышается тонус матки, стимулируются частота и амплитуда схваток.
Серотонин оказывает выраженное влияние на состояние и функцию клеток миометрия, угнетает активность холинэстеразы и усиливает действие ацетилхолина, способствует передаче возбуждения с двигательного нерва на мышечное волокно.
Действие серотонина на матку может осуществляться двумя путями: непосредственно на миометрии через a-рецепторы и через центральную нервную систему путем усиления выработки окситоцина нейрогипофизом.
Кинины усиливают сократительную способность матки за счет увеличения скорости кровотока в ней.
Определенное значение в развитии родовой деятельности имеет гистамин, способствующий выработке гипофизом окситотических веществ.
Существует предположение, что изменения гормональных взаимоотношений у матери перед родами тесно связаны со степенью зрелости гормональной регуляции у плода, которая генетически детерминирована завершенностью процессов его роста и развития. В первую очередь важное значение имеет зрелость эпифиз-гипоталамо-гипофизарной системы плода.
Влияние гормонов плода на подготовку и начало родов складывается из следующих моментов:
1. при повышении уровня плодового кортикотропина в крови матери увеличивается содержание эстрадиола и уменьшается количество прогестерона, хориогонадотропина и хориомаммотропина (плацентарного лактогена);
2. выделяющийся перед родами у плода окситоцин действует аналогично окситоцину матери;
3. перед родами уровень мелатонина в крови у плода повышается, а у матери – понижается. Указанные изменения концентрации мелатонина способствуют перестройке стероидного гемостаза и формированию эстрогенного фона в организме беременной. Мелатонин изменяет соотношение гтролактин/фоли/лютропин в сторону увеличения двух последних. В результате происходит повышение синтеза эстриола. Наряду с этим снижение уровня мелатонина в крови матери приводит к освобождению лейкотриенов (медленно реагирующих субстанций анафилаксии), что в свою очередь ведет к активации кейлонной активности и торможению дальнейшего увеличения массы плода, его органов и тканей и способствует активации механизмов подготовки организма беременной к родам. Одновременное подавление синтеза иммунодепрессантов пролактина и хориогонадотропина приводит к усилению трансплантационного иммунитета и стимулирует отторжение плода как аллотрансплантата;
4. ишемия оболочек плодного яйца перед родами, обусловленная повышением внутри маточного давления, активизирует "арахидоновый каскад" с выделением утеротонических простагландинов.
Наряду с изменениями в нервной и гуморальной регуляции перед родами существенные изменения происходят в репродуктивной системе, в том числе в матке.
В основе сокращения мышц матки, как и других органов (сердца, сосудов), лежит процесс превращения химической энергии в механическую.
Особое значение для начала родов имеют следующие процессы, происходящие в матке:
1. увеличение в миометрий интенсивности метаболических процессов, скорости потребления кислорода (в 3,5 раза), содержания сократительного белка актомиозина (на 25 %) , гликогена, глутатиона, фосфорных соединений (АТФ, креатинфосфат, фосфокреатинин), играющих большую роль в энергетических процессах мышечной ткани;
2. изменения соотношения между мышечной и соединительной тканью в сторону резкого преобладания первой из них, особенно в теле матки.
В результате изменений, происходящих в мышечных клетках, снижается мембранный потенциал, увеличивается их возбудимость, усиливается спонтанная активность, повышается чувствительность к контрактильным веществам.
С началом родов в матке формируется группа клеток, в которых первично возникает возбуждение, распространяющееся в последующем на всю матку. Эта область называется водителем ритма ("пейсмекер"). Она располагается в дне матки, ближе к правому углу.
Изменения соотношения гормонов и биологически активных веществ, влияющих на возбудимость и сократительную деятельность матки перед родами, проходит в несколько этапов: первый этап характеризуется состоянием зрелости гормональной регуляции плода; второй этап – активацией эстрогенов и изменениями в матке; третий этап – синтезом утеротонических соединений, в первую очередь простагландинов, окситоцина – основных факторов, обеспечивающих развитие родовой деятельности.
Важное значение имеют не только выброс утеротонических соединений перед родами, но и пульсирующий тип их синтеза во время родов, что обеспечивает регулярный характер родовой деятельности.
В результате изменений, происходящих в нервной и гуморальной регуляции, а также в самой матке, формируется попеременное возбуждение центров симпатической и парасимпатической иннервации:
1. под влиянием медиаторов симпатической нервной системы (норадреналин и адреналин) происходит сокращение продольно расположенных мышечных пучков в теле матки миометрия при одновременном активном расслаблении циркулярно (поперечно) расположенных пучков в нижнем сегменте;
2. в ответ на максимальное возбуждение центра симпатической нервной системы и выделение большого количества норадреналина происходит возбуждение центра парасимпатической нервной системы;
3. под влиянием медиатора парасимпатической нервной системы (ацетилхолин) сокращаются циркулярные мышцы при одновременном расслаблении продольных;
4. по достижении максимального сокращения циркулярных мышц наступает максимальное расслабление продольных;
5. после сокращения матки наступает период полного ее расслабления (пауза между схватками), когда происходит восстановительный синтез сократительных белков миометрия.






